



Ингибиторы биосинтеза ароматических аминокислот
В качестве представителя данного класса гербицидов рассмотрим глифосат — неселективный гербицид широкого спектра действия, открытый в 1971 г. Практически он не токсичен для млекопитающих, птиц, рыб, насекомых и большинства бактерий. Гербицид легко разлагается в почве на аммиак, CO2 и неорганический фосфат. Глифосат — первый коммерчески значимый гербицид, чья мишень представляет собой фермент 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS), катализирующий предпоследнюю реакцию шикиматного пути в растениях.
Как системный гербицид глифосат передвигается в растении по флоэме и накапливается в акцепторных зонах, включая точки роста, зоны аккумуляции углерода и высокой метаболической активности. Действие глифосата начинается через несколько минут после обработки и продолжается дни и недели. Индикаторами токсичности гербицида являются заторможенный рост, хлороз и опадание листьев, а также некроз тканей. Кроме ингибирования биосинтеза ароматических аминокислот, гербицид вызывает, в частности у растений сахарной свеклы, нарушения синтеза белка и фотосинтетического метаболизма углерода.
Широкая картина действий глифосата на органы растений и процессы представлена в таблице 14.2.
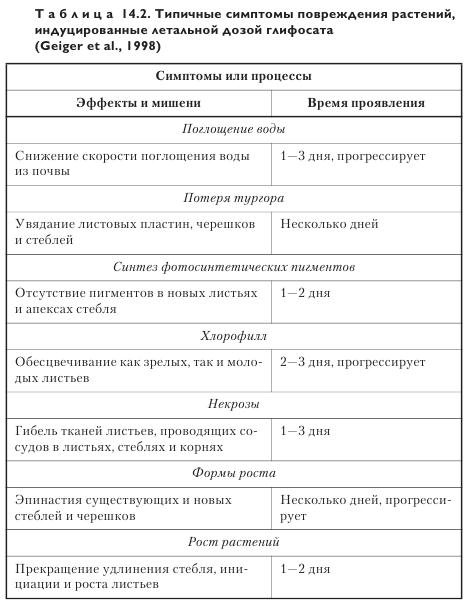

Симптомы повреждения, индуцированные глифосатом, проявляются в определенной последовательности, начиная с постепенного развития видимых симптомов, таких, как приостановка роста и пожелтение тканей. В последующем происходит изменение окрашивания побегов и корней вплоть до коричневого, на что требуется от нескольких дней до недель. Время и скорость проявления специфичных симптомов зависит от вида растения, возраста, фазы развития, окружающих условий и концентрации глифосата.
Так, различия в симптомах повреждений обнаружены между сахарной свеклой, высокочувствительной к глифосату, и канатником — проблемным сорным растением этой культуры, более толерантным к данному гербициду. Хотя схема его первичного действия заключается в ингибировании EPSPS, превращающем фосфоенолпируват (ФЕП) в шикимат-3-фосфат, и одинакова для обоих видов, что подтверждается накоплением шикимата, контрастный физиологический ответ на обработку гербицидом приводит к заметным различиям между культурными и сорными растениями в развитии симптомов повреждения.
В отличие от контактных гербицидов, действующих локально, глифосат оказывает эффект не только в зоне внесения, но и в местах его транспортировки. Проходя через кутикулу и плазмалемму, глифосат оказывается во флоэме и транспортируется по ней вместе с сахарозой к акцепторам, накапливаясь в цитозоле, в вакуолях и строме хлоропластов. После поступления в пластиды гербицид блокирует шикиматный путь, ингибируя фермент EPSPS. Шикиматный путь состоит из семи биохимических реакций и завершается образованием хоризмата, который, в свою очередь, через серию побочных реакций превращается в ключевые ароматические аминокислоты — фенилаланин, тирозин и триптофан. Главные причины повреждения растений, возникающие из-за ингибирования EPSPS: 1) блокирование синтеза хоризмата, влияющего на различные аспекты метаболизма, связанные с продуктами шикиматного пути; 2) дезинтеграция цикла фотосинтетического восстановления углерода, оказывающая воздействие на фотосинтез и регуляцию метаболизма углеводов. Их взаимодействие и определяет значительное разнообразие летальных эффектов от применение глифосата.
Влияние глифосата на целый ряд важнейших физиологических процессов в растении и возможные механизмы этого влияния показаны в таблице 14.3 и на рисунке 14.1.


Быстрота и глубина изменения процессов зависит от вида и биотипа, что указывает на то, что механизмы летальных эффектов тоже могут различаться. Если, например, у данного биотипа или вида глифосат одновременно вызывает быстрое снижение фотосинтеза и содержания интермедиатов (ФГК), можно предположить, что симптомы повреждения связаны с рассогласованием поступления углерода в шикиматный путь. Снижение содержания ФГК в листьях сахарной свеклы увеличивает соотношение Рн/ФГК, ингибирует активность ADPG-пирофосфорилазы и, соответственно, синтез крахмала. Ингибирование фотосинтеза замедляет синтез и транспорт сахарозы. Если подавление фотосинтеза сильное, что часто встречается у сахарной свеклы, происходит фотоингибирование, что особенно заметно на второй день, когда ИФ начинает снижаться по мере приближения освещенности к полуденному максимуму.
Порядок появления и динамику развития симптомов также можно использовать для выявления причины нарушения физиологических процессов (прямое действие глифосата или последствия ухудшенного состояния растений). Интересно, что влияние обработки гербицидом 12-дневного растения пшеницы сказалось на транспирации (через несколько часов), фотосинтезе (через 3 дня) и дыхании (через 9 сут). При этом влияния глифосата на отдельные фазы фотосинтеза не установлено, т. е. фотосинтез не является первичной мишенью действия глифосата.

Несмотря на то что глифосат детален для большинства растений, динамика и интенсивность развития симптомов неодинаковы у разных видов. При этом неясно, какой именно процесс нарушается у тех или иных видов. Так, нарушения в акцепторных зонах корней могут лимитировать поглощение воды и питательных веществ, а в донорных листьях — постоянное поступление ассимилированного углерода в обмен веществ. В здоровых растениях поступление углерода в донорный лист, а воды и минеральных веществ в корни поддерживают баланс питательных веществ, значимый для жизни растений. Нарушения в цикле фотовосстановления углерода в донорном листе сахарной свеклы — главный фактор летального действия глифосата, ведущий к ухудшению обеспечения ассимилятами тканей акцептора (табл. 14.4).


У сорного растения канатника нарушения фотосинтеза выражены в меньшей мере, а главный летальный фактор — нарушение метаболизма в акцепторе. У обоих видов после действия глифосата серьезно нарушается поглощение воды, хотя причины этого разные. Проблемы доступности углерода резко лимитируют у сахарной свеклы целостность тканей как донора, так и акцептора, в то время как у канатника непрерывный экспорт и накопление глифосата нарушают целостность клеток корня. Видимо, именно быстрое и полное подавление фотосинтеза приводит к большей чувствительности к глифосату сахарной свеклы, в то время как относительно медленное ингибирование фотосинтеза у канатника является предпосылкой большей его толерантности.
Благодаря генетической трансформации растений разными формами EPSPS, изменяющими первичный способ действия глифосата, на некоторых культурах достигнута устойчивость к глифосату. Избраны три стратегии генетической трансформации:
• синтез избыточного количества EPSPS;
• введение гена, вызывающего распад гербицида;
• внедрение EPSPS с пониженным сродством к глифосату.
Наиболее эффективной оказалась последняя стратегия, основанная на воспрепятствовании присоединения глифосата к EPSPS. В результате длительных генно-инженерных работ создан устойчивый к глифосату сорт сахарной свеклы, содержащий ген EPSPS, который был получен из почвенных бактерий Agrobacterium. Эта форма в полевых дозах глифосата не обнаруживает симптомов повреждения ни шикиматного пути, ни фотовосстановления углерода. В результате был достигнут такой уровень резистентности, что для 50%-го подавления активности EPSPS концентрацию глифосата нужно было увеличить в 500 раз.
Из Е. coli клонирован ген, кодирующий слабочувствительную к глифосату EPSP-синтазу. Введение этого гена в табак привело к получению устойчивых к глифосату растений. Полученные трансгенные растения Petunia hybrida с высоким уровнем экспрессии гена аrоА были также устойчивыми к глифосату. Линия сои с агро-бактериальным геном, слабо чувствительным к глифосату, была очень устойчива к гербициду в полевых условиях, перенося обработку глифосатом в дозе до 1,68 кг/га без видимых повреждений. Клонирован также ген глифосатоксидоредуктазы (GOX), метаболизирующий глифосат в нетоксичное соединение — аминометилфосфоновую кислоту. Комбинация СР4- и GOX-генов использована фирмой Монсанто в качестве селектируемых при трансформации кукурузы и некоторых двудольных. Получены трансгенные растения пшеницы, устойчивые к коммерческим концентрациям глифосата.
Однако, несмотря на то что трансгенные растения, экспрессирующие гены EPSPS, демонстрировали высокие уровни толерантности, полевые испытания показали, что в действительности толерантность к глифосату повысилась в 2 раза (растения переносили в 2 раза большую концентрацию глифосата по сравнению с рекомендуемой). Этого пока недостаточно для коммерческого использования данного метода.
В литературе есть данные о наличии биотипов вьюнка полевого (Convulvulus awensis), повысивших устойчивость до 4 раз, но не различающихся ни по суммарному поглощению гербицида, ни по его распределению по органам. Видимо, различия по толерантности обусловлены уже на клеточном/метаболическом уровне. Отдельные особи в популяции многолетнего райграса также демонстрируют высокий уровень толерантности, достигнутый, в частности, в результате одиннадцатилетней возвратной селекции. Высокая степень толерантности отмечена для Lolium rigidum — ценной пастбищой травы, являющейся также сорным растением в посевах зерновых в Южной Австралии. Популяция высокоустойчивых биотипов обнаружена на полях, где глифосат вносился 10 раз (за последние 15 лет) для борьбы с сорняками перед посевом. Устойчивость этих биотипов была в 10 раз выше, чем у восприимчивых биотипов. Этим, возможно, объясняется индукция глифосатом нескольких ферментов, разрушающих сам гербицид. Вероятно, имеются и другие причины, как, например, обнаруженные при сравнении толерантных и восприимчивых форм Lolium rigidum различия по доступу глифосата к ферменту EPSPS. По большинству других параметров различий нет. Видимо, именно поэтому устойчивые биотипы каким-то образом обходили влияние вторичных нарушений на физиологические процессы и развитие симптомов повреждения. Сниженная скорость транспорта гербицида к месту действия в пластидах, видимо, тоже может сказаться на толерантности, наряду с различной активностью фосфат/фосфат- и ФЕП/фосфат-транслокаторов, расположенных во внутренней мебране хлоропластов. Они регулируют обмен метаболитами между цитоплазмой и хлоропластами и могут ограничивать поступление гербицида в строму пластиды. Подобный механизм подтверждает высказанную выше гипотезу, что толерантность райграса определяется на клеточном/метаболическом уровне.
Специфической проблемой использования генетически модифицированных культур, устойчивых к гербицидам, считается предотвращение возможности переноса генов устойчивости к сорным растениям путем переопыления с дикими родственниками. В этих целях применяют генетическую трансформацию ДНК органелл (митохондрий, пластид), поскольку они наследуются по материнской линии и не могут быть переданы с пыльцой диким родственникам. Так, успешно проведена встройка в пластиды табака гена EPSPS петунии, который обеспечивает устойчивость к глифосату. Трансгенные растения сорго, устойчивые к гербицидам, получали бомбардировкой незрелых эмбрионов на стадии зиготы микрочастицами золота (диаметр частиц 1,5—3 мкм). Микрочастицы погружали в раствор экспрессирующего вектора, высушивали и «выстреливали» в клетки-мишени, добиваясь при этом высоких результатов трансфекции.