



Реакция растений на повышение концентрации углекислого газа в атмосфере
Адаптация растений к повышению концентрации углекислого газа. Повышение концентрации CO2 в атмосфере почти на 28% за последние 250 лет (рис. 13.2) вызывает естественный вопрос — в какой мере это воздействие является стрессовым для растении в по-левых условиях?
Исследования на гербарном материале показывают, что подобная реадаптация может вызвать значительное (до 70%) снижение плотности устьиц, причем повышение концентрации CO2 оказывает влияние на инициацию устьиц, а не просто на растяжение эпидермальных клеток. Таким образом, повышение концентрации CO2 отрицательно коррелирует с плотностью устьиц на фоне меньшей дискриминации 13C (A13С) в листьях, что предполагает повышение ЭИВ. Одновременно за последние 250 лет снизилось содержание питательных веществ в листьях, включая соотношение N/C. Однако отмеченная тенденция может быть связана кроме повышения концентрации CO2 и с другими антропогенными воздействиями на условия роста и почвенное плодородие.
Двуокись углерода является прежде всего субстратом фотосинтеза и действия ключевого фермента РБФК/о у С3-растений, составляющих 95% всех растений (рис. 13.3).


У полевых культур с C3- и С4-типом фотосинтеза KМ (CO2) составляет соответственно 10 и 30 мкмолей. РБФК характеризуется также оксигеназной активностью, что реализуется в процессе фотодыхания. Современное соотношение СО2/О2 в атмосфере вызывает почти 40%-е снижение фотосинтеза у С3-видов при 25 °C, причем дальнейшее повышение температуры усиливает ингибирование. Благодаря конкурентному взаимодействию удвоение концентрации CO2 в атмосфере уменьшает ингибиторный эффект O2 наполовину, одновременно снижая темновое дыхание, хотя и не у всех растений. Указанное снижение имеет две компоненты, причем одна из них связана с непосредственным ингибированием дыхательного метаболизма, прекращающимся сразу же после снижения концентрации CO2. Вторая компонента представляет собой более продолжительное снижение ИД. В некоторых случаях это может быть следствием ослабления дыхания в результате снижения содержания азотистых соединений, поддержание уровня которых более энергоемко, чем, например, углеводов. Поэтому ИД единицы N биомассы, как правило, зависит от концентрации CO2.
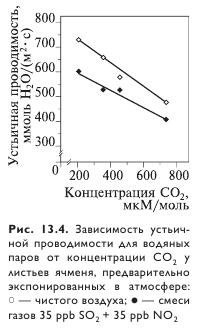
Устьичная проводимость при повышенной концентрации CO2 также снижается (рис. 13.4). Однако, поскольку одновременно повышается площадь листьев, расход воды в расчете на одно растение может и не уменьшиться. Эффективность использования воды, напротив, обычно увеличивается в результате ассимиляции большего количества углерода.
Кроме того, повышение концентрации CO2 является своего рода модулятором ростовых процессов, приводя к изменениям в анатомии, морфологии и фенологии, и косвенно воздействует на фотосинтез посредством изменения различных характеристик растения, включая ветвление, площадь листьев, продолжительность ассимиляции CO2 и мощность акцептора.
Разнообразие фотосинтетических реакций на обогащение атмосферы углекислым газом. Одним из хорошо изученных феноменов адаптации является зависимость ИФ от внутриклеточной концентрации CO2 или А/сi кривая, имеющая характер гиперболы. В нижней части кривой фотосинтез лимитируется количеством РБФК, а на плато — скоростью регенерации РБФ. Во время фотосинтеза листья С3-видов пытаются поддерживать ci в зоне перехода от линейной части кривой к плато таким образом, чтобы как РБФК, так и скорость регенерации РБФ совместно лимитировали ИФ и поддерживали величину соотношения c/ca1 около 0,7. При удвоении концентрации CO2 устьичная проводимость снижается, но соотношение ci/ca не меняется, что указывает на отсутствие независимой устьичной адаптации (рис. 13.5). При этом устьичное лимитирование снижается, так как ci повышается с 245 до 490 мкмоль/моль. Повышение ci и вызывает начальное 50%-ное увеличение ИФ, что отмечается во многих опытах; это сдвигает фотосинтез на кривой A/ci в зону лимитирования рибулозобифосфатом.

Во время длительного воздействия повышенной концентрации CO2 на растения отмечены разные типы адаптации фотосинтеза у полевых культур на уровне кривой А/ci, выражающиеся в изменениях угла наклона линейной части и/или в зоне лимитирования РБФ. Так, у сои выше как угол наклона, так и уровень плато фотосинтеза, в то время как у хлопчатника изменений нет. У капусты оба этих показателя оказались даже ниже, чем при атмосферной концентрации CO2.
Маленький объем сосуда или дефицит питания, как правило, снижают ИФ(А) при данной ci, хотя такая тенденция может и не подтвердиться в полевых условиях, поскольку при этом снижается адаптация на уровне кривой A/ci. Низкая температура, конкуренция за свет, воду и питание могут ограничивать эффект повышенной концентрации CO2.
В большинстве случаев повышение ассимиляции CO2 при повышенной его концентрации, вероятно, указывает на ограниченную способность использовать избыточное количество углерода (схема регуляции фотосинтеза по принципу обратной связи) из-за недостаточности других факторов среды или из-за генетически детерминированных ограничений. Тогда адаптация фотосинтеза представляет собой скорее процесс оптимизации, нежели реакцию на повышение концентрации CO2 как на стресс. Она включает перераспределение ресурсов от нелимитирующих звеньев, таких, как доступность углерода, в более лимитирующие звенья — поглощение квантов света, транспорт электронов, утилизация углеводов.
РБФК играет ключевую роль в адаптации, причем ее активность у целого ряда видов (культур) при повышенной концентрации CO2 ниже, чем в нормальных условиях. Иногда содержание белка РБФК падает на 60%, что указывает на перераспределение азота и является одной из причин снижения активности РБФК. Интересно, что снижение содержания белка фермента может происходить и в присутствии достаточного количества азота в среде.
Однако наиболее частым случаем адаптации и снижения активности РБФК является нарушение баланса в донорно-акцепторных отношениях. У трансгенных растений сверхэкспрессирование кислой инвертазы и, соответственно, избыточное накопление гексоз снижало фотосинтез, ферментативную активность и количество РБФК. Глюкоза представляет собой регуляторный сигнал, репрессирующий транскрипцию фотосинтетических генов, включая кодирующие малую и большую субъединицы РБФК — rbc S и rbc L.
Кроме того, гены, непосредственно связанные с метаболизмом углеводов, могут положительно или отрицательно регулироваться сахарами. Такая регуляция может быть средством повышения активности ферментов трансформации углеводов, а также баланса между донором и акцептором. Так, у риса, выращиваемого при повышенной концентрации CO2, повышение содержания сахарозы и крахмала коррелирует с 20%-м увеличением активности сахарозо-фосфатсинтазы (СФС) при одновременном снижении активности и содержания РБФК. Однако есть и другие данные, в частности полученные на фасоли, когда активность СФС при повышенной концентрации CO2 ниже, чем при нормальной.
Выращивание С3-растений при повышенной концентрации CO2 снижает не только количество углерода, вступающего в цикл Кальвина, но и потоки азота в фотодыхательном цикле, которые почти на порядок выше количества поглощаемого корнями азота. Обычно концентрация азота в листе, как и активность нитратредуктазы и содержание нитратов, ниже при повышенных концентрациях CO2. Видимо, нитраты могут выступать в качестве метаболического сигнала для протеинкиназ при регуляции потоков углерода между сахарозой и аминокислотами. Если это действительно так, уменьшение содержания нитратов в листе при повышенной концентрации CO2 в листе может перенаправить углерод от аминокислот на синтез сахарозы по механизму, аналогичному репрессии глюкозой экспрессии генов.
Что касается реакции С4-растений на повышенную концентрацию CO2, она, как правило, позитивная, хотя и не столь заметная, как у С3-видов. Однако встречаются случаи, когда реакция отсутствует или даже отрицательна. Есть несколько причин стимулирующего эффекта повышенной концентрации CO2 на С4-растения. Во-первых, это уменьшение (как и у С3-растений) устьичной проводимости, в результате которого ЭИВ может повышаться на 50%. Это, в свою очередь, может стимулировать ростовые процессы даже в условиях ограниченной водообеспеченности. Во-вторых, повышенная концентрация CO2 стимулирует кущение (побегообразование) и формирование листьев. В результате суммарная ассимиляция CO2 растением оказывается выше, несмотря на пониженную ИФ единицы площади листа. В-третьих, недостаточное азотное питание и высокая засоленность усиливают выход CO2 из клеток обкладки проводящих пучков, увеличивая чувствительность С4-растений к повышению концентрации CO2.
В плотных (загущенных) посевах в безветренные дни и, соответственно, при слабом турбулентном перемещивании воздуха са может быть достаточно низкой, что сдвигает С4-фотосинтез в линейную часть кривой A/ci. В этом случае фотосинтез также будет положительно реагировать на повышение концентрации CO2.
Кратковременное повышение концентрации CO2 в атмосфере, как правило, вначале увеличивает ИФ. Синтезирующиеся неструктурные углеводы при этом накапливаются в листьях и других органах растений в форме крахмала и/или растворимых углеводов (в зависимости от вида) (рис. 13.6). В некоторых случаях возможно ингибирование фотосинтеза по принципу обратной связи из-за накопления повышенных концентраций крахмала, что может свидетельствовать о лимитировании процессов роста недостаточной мощностью акцептора или проблемах загрузки флоэмы и транспорта растворимых углеводов.

Это может привести к постепенному торможению фотосинтеза, хотя и не во всех случаях, как, например, у сои, у которой ИФ даже повышается. Листья сои часто формируют дополнительный слой клеток мезофилла, в стеблях и листьях накапливается больше структурных углеводов. Соя, в отличие, например, от пшеницы (вид с С3-типом метаболизма), обладает возможностью:
• симбиотического связывания N2;
• формирования дополнительных слоев палисадных клеток в листовой паренхиме;
• перераспределения ассимилированного при фотосинтезе углерода в большей степени в относительно инертный крахмал, чем в растворимые сахара;
• создавать мощные акцепторы ассимилятов в виде листьев и стеблей в фазе вегетативного развития и наливающихся семян в фазе генеративного развития.
Виды, у которых отсутствуют эти характерные признаки, вряд ли могут избежать ингибирования фотосинтеза, по меньшей мере частичного, при повышенной концентрации CO2 в атмосфере.
Интересно, что избыточное количество ассимилятов, синтезируемое при повышенном количестве CO2, как правило, направляется к корням, а не в побеги, где больше ассимилятов поступает в стебли и поддерживающие структуры, чем в листья.
Что касается конкурентных взаимоотношений C3- и С4-видов в ценозах и естественных экосистемах, они не так однозначны, как можно было бы ожидать на основании только реакций фотосинтеза на повышенную концентрацию CO2. Другие факторы среды также оказывают заметное влияние на исход конкуренции, причем концентрация CO2 при этом может и не быть главным лимитирующим фактором. Важно, что отдельные растения и растения в ценозе по-разному реагируют на фактор CO2. Среди С3-видов также есть генотипические различия по реакции на CO2, что осложняет оценку результатов конкурентных взаимоотношений, хотя по большинству прогнозов С3-видам отдается больше предпочтений.
Опыты 1970-х гг. с 30 C3- и С4-видами показали стимуляцию накопления биомассы при повышенной концентрации у C3-, но не у С4-видов. Так, у 8 основных С3-культур удвоение концентрации CO2 вначале увеличивало нетто-фотосинтез в среднем на 52%, впоследствии он снижался до 29%. Тем не менее биомасса и продуктивность опытных растений оказались в среднем на 30 и 41% выше, чем в контроле. Длительное выращивание при повышенной концентрации CO2 затронуло кроме биохимии фотосинтеза и физиологии устьиц адаптацию на уровне площади листьев, ИЛП, ФСП, толщины листовой пластинки, степени ветвления и кущения, биомассы стеблей и корней, размера плодоэлементов, фенологию. Интересно, что, если даже ИФ листа снижается, изменения других параметров, в частности ФСП и ИЛП, приводят к формированию большей биомассы и соответственно повышению урожайности.
Влияние факторов среды на реакцию растений на повышенную концентрацию углекислого газа. Повышенная концентрация CO2 как парникового газа вызывает также повышение температуры воздуха. Температура и углекислый газ характеризуются интерактивным эффектом, поскольку повышение температуры снижает соотношение [СO2]/[O2] в растворе, повышает интенсивность фото- и темнового дыхания, а также аттрагирующую способность акцептора. Расчеты на биохимической модели показывают, что в случае отсутствия ингибирования активности РБФК и удвоения концентрации CO2 до 650 мкмоль/моль ИФ при световом насыщении могла бы повыситься при 25 °C на 20%, а при 35 °C на 105% при одновременном повышении температурного оптимума на 5°С. Даже при 40%-ной потере активности РБФК нетто-фотосинтез все еще мог бы быть выше при температуре, превышающей 25 °C.
Этот выигрыш в ассимиляции углерода может реализоваться в более высокой продуктивности, но может и не реализоваться из-за взаимодействия факторов, усложняющих ситуацию. Так, например, листья реагируют на повышенную температуру воздуха более активной транспирацией, в то время как увеличение концентрации CO2 повышает температуру листа из-за снижения транспирации. Взаимодействие температуры и CO2 может оказать больший интерактивный эффект на формирование площади листьев, чем непосредственно на фотосинтез. При этом следует иметь в виду, что температуры, стимулирующие при повышенной концентрации CO2 вегетативный рост, могут оказывать негативное влияние на формирование урожая. Так, зерновая продуктивность риса, сои и пшеницы при обогащении атмосферы CO2 и температуре выше 26 °C снижалась примерно на 10% на каждый градус. Таким образом, фазы вегетативного и репродуктивного развития отражают интегральную реакцию метаболизма и онтогенетических процессов на температуру. В итоге видовые особенности, фазы развития, световой режим, условия минерального питания, температурный режим — все они модифицируют реакцию растений на характер связи между температурой и CO2.
Относительное стимулирование фотосинтеза и вегетативного роста высокими концентрациями CO2 усиливается с повышением температуры в некоторой степени даже у С4-видов, доходя в среднем до 2% на 1 °C. Характер взаимосвязи между концентрацией CO2 и температурой видоспецифичен. Так, соотношение биомассы при повышенной и нормальной концентрации CO2 при 25 °C у редиса и моркови составило соответственно 1,5 и 2, а у подсолнечника и кормовых бобов, напротив, повышенные температуры оказывали отрицательное влияние на продуктивность.
Опыты с 20 C3- и 9 С4-видами показали, что удвоение концентрации CO2 (по сравнению с естественным содержанием) удваивает АТО, определяемое как соотношение интенсивностей фотосинтеза и транспирации. Различия между двумя группами видов статистически недостоверны, хотя при естественной концентрации CO2 ATO у С4-видов было на 50% выше. Повышение ATO при обогащении воздуха CO2 объясняется повышением ИФ и снижением ИТ, причем последнее особенно важно в условиях дефицита влаги в корнеобитаемой среде. Однако этот эффект может полностью исчезать, если повышение концентрации CO2 происходит на фоне увеличения температуры воздуха.
Что касается продуктивности транспирации (как показателя эффективности использования воды), при удвоении концентрации CO2 с 350 до 700 мкм/моль она увеличилась у фасоли (С3-вид) с 3 до 4,4 г сухой массы/кг H2O, а у сорго (С4-вид) с 4,2 до 4,9 г сухой массы/кг H2O. Таким образом, ПТ у С3-видов увеличивается сильнее, чем у С4-видов.
Для большинства культур повышение концентрации CO2 снижает ингибиторный эффект умеренного солевого стресса. В этом опыте концентрация солей в листьях опытного и контрольного вариантов примерно одинакова, что указывает на четкую регуляцию транспорта солей, несмотря на более активный рост и меньшую скорость водопотребления при повышенной концентрации CO2.
Повышение концентрации CO2 в условиях недостаточного минерального питания обычно вызывает относительную стимуляцию фотосинтеза и роста, хотя дефицит питания, как правило, подавляет ИФ. Причина в том, что концентрация элементов питания при обогащении воздуха CO2 может снизиться на 20%, хотя их общее содержание на одно растение может даже увеличиться из-за накопления большей биомассы. Содержание азота в расчете на одно растение, например, увеличивается, но не в такой степени, как углерода. Скорость же поглощения азота даже снижается из-за уменьшения интенсивности транспирации. В любом случае концентрация азота при обогащении воздуха CO2 уменьшается, а интенсивность использования азота возрастает даже на высоком фоне азотного питания. Концентрация фосфора в тканях, напротив, мало зависит от содержания CO2 в атмосфере.
Что касается освещенности, то повышение концентрации CO2 увеличивает ИФ на плато фотосинтеза. В результате снижения кислородного ингибирования увеличивается квантовый выход фотосинтеза. При сравнении световых кривых фотосинтеза С3-растений, выращиваемых при концентрации CO2 330 и 660 мМ/моль, в последнем случае отмечаются более высокий уровень плато фотосинтеза и больший угол наклона линейной части, но световой компенсационный пункт при этом ниже. Благодаря повышению квантового выхода фотосинтеза, фотосинтез посева, в частности сои, увеличивался линейно с повышением концентрации CO2 со 160 до 990 мМ/моль даже после смыкания рядков в междурядьях и достижения максимального поглощения квантов света посевами. Повышение квантового выхода имеет важные последствия для посевов, увеличивая фотосинтез в облачные дни почти на 90%.
Таким образом, повышение концентрации CO2 в воздухе системно влияет на характер взаимосвязи всех физиологических процессов в растении. Сложность этих связей для анализа особенно возрастает на фоне изменения других климатических параметров, в частности температуры воздуха и влагообеспеченности. При этом представляют особый интерес видоспецифические механизмы адаптации к сочетанию указанных и иных стрессоров. Рассмотрим это подробнее на примере сои, риса и пшеницы.
Соя. Фотосинтез. При длительном выращивании при удвоенной концентрации CO2 (330 —> 660 мкМ/моль) квантовый выход фотосинтеза у растений сои возрастал с 0,05 до 0,09, максимальный фотосинтез (при насыщающей освещенности) составлял 51,8 и 126,6 мкМ С02/(м2*с), константа Михаэлиса (KМ) — 359 и 1133 мкм/моль, углекислотный компенсационный пункт (УКП) — 63 и 42 мкМ/моль соответственно. Интересные данные получены при определении активности и количества РБФК в листьях растений, выращиваемых при концентрациях CO2 160, 220, 280, 330, 660, 990 мкМ/моль. Активность фермента была практически постоянной при разных режимах выращивания и составила 1,0 мкМ С02/(мин*мг) растворимого белка. Содержание белка не зависело от концентрации CO2 при выращивании и не превышало 2,4 г/м2 листьев, причем 55% белка принадлежало фракции I. УПП листьев увеличивалась по мере повышения концентрации CO2, в результате чего активность РБФК в расчете на единицу площади листа уменьшалась. В аналогичных условиях выращивания доля белка РБФК у сои (по сравнению с рисом) не зависела от концентрации CO2 и составила в среднем около 55% (табл. 13.5).

Дыхание. ИД посевов сои в фазу вегетативного развития при выращивании при 320 и 640 мкМ/моль CO2 составила соответственно 2—3 и 4—5 мкМ СО2/(м2*с), причем утренние значения ИД посевов после перехода растений с одного варианта на другой быстро адаптировались к новому режиму выращивания. Удельная ИД (в расчете на единицу азота) снижалась как при кратковременном действии, так и при длительном выращивании при повышенной концентрации CO2.
Эффективность использования воды. При обогащении воздуха CO2 ИТ снижается, а ИФ, напротив, увеличивается, что приводит к повышению ATO и ЭИВ. Увеличение последней обеспечивается на 80% возрастанием ИФ и на 20% снижением ИТ.
Углеводы. Соя накапливает неструктурные углеводы, особенно крахмал, при обогащении воздуха CO2. При выращивании при 330, 450, 600 и 800 мкМ/моль CO2 содержание сахарозы, редуцирующих сахаров в листьях в течение дня остается примерно одинаковым в изучаемых вариантах, а содержание крахмала постоянно возрастает со скоростью примерно 0,006% за 1 ч (в расчете на сухую массу). В диапазоне концентраций CO2 330—800 мкМ/моль общее содержание сахаров увеличилось с 0,002 до 0,004%, а крахмала с 0,085 до 0,204% (в расчете на сухую массу). Подобная тенденция отмечена во все остальные фазы развития.
Азот. Содержание азота в сухой массе растения по мере увеличения CO2 снижалось с 0,050 до 0,037%. При пересчете содержания азота на структурную массу (за вычетом неструктурных углеводов) разница между крайними вариантами сглаживалась и составила при минимальной и максимальной концентрации CO2 соответственно 0,055 и 0,048%.
Скорость роста. В фазе вегетативного развития после полного смыкания рядков в посеве скорость роста варьировала в пределах 5—20,7 г/(м2*сут) при почти 6-кратной (160—990 мкМ/моль) разнице в концентрациях CO2 при выращивании. Сухая биомасса растения в крайних вариантах при уборке составляла соответственно 12,9 и 39,1 г, а масса семян — 5,8 и 17,8 г.
Взаимодействие температуры и диоксида углерода. Повышение концентрации CO2 при выращивании всегда увеличивало накопление биомассы и урожайность, но снижало Kхоз. Эффект углекислотной подкормки растений, как правило, возрастает с повышением температуры воздуха до определенного уровня на фоне сокращения периода налива семян. Однако урожайность семян по мере повышения дневной (ночной) температуры имела тенденцию к некоторому снижению (табл. 13.6.).

Число семян на растении с повышением температуры выращивания и концентрации CO2 в климакамерах несколько повышалось. Масса единичного семени при разных вариантах концентрации CO2 снижалась при повышении температуры, как и Kхоз, хотя семенная продуктивность и масса побега при обогащении атмосферы CO2 возрастали. Согласно данным других опытов, семенная продуктивность резко снижалась при температуре 40/30 °C (день/ ночь) и выше, биомасса начинала быстро снижаться при температурах 44/34 °C и выше. Снижение семенной продуктивности сои при повышенной температуре выращивания в опытах объясняется сокращением периода налива семян. Селекционерам в будущем придется вводить в зародышевую плазму признаки устойчивости к высокой температуре на фоне модифицированной фотопериодической реакции. Кроме того, должна быть создана зародышевая плазма, лучше использующая избыточные количества фотоассимилятов, образующихся при обогащении атмосферы CO2.
Рис. Фотосинтез. ИФ листьев риса, выращиваемого при трех температурных режимах (день/ночь) — 32/23, 35/26 и 38/29 °C увеличилась при удвоении концентрации углекислого газа с 330 до 660 мкМ/моль в среднем по всем температурам с 18,8 до 30,4 мкМ СО2/(м2*с), т. е. на 60%. Интересно, что кривые зависимости ИФ от внутриклеточной концентрации CO2 при обеих концентрациях CO2 хорошо совпадают, что характерно для всех температур (см. рис. 13.5). Экстраполяция линейной части углекислотной кривой до оси абсцисс дает величину углекислотного компенсационного пункта (УКП) в листе около 60 мкМ/моль CO2.
Нетто-фотосинтез посева после смыкания рядков (в отличие от ИФ отдельных листьев) возрастает линейно с увеличением мощности потока квантов света, не достигая насыщения на полном солнечном свету. Это обусловлено эректоидностью листьев и высокой густотой посева. Так, ИФ посева при выращивании при 160, 250, 330, 500, 660 и 900 мкМ/моль CO2 варьировала от 34 до 90 мкМ CO2/(м2*с), т. е. при почти 6-кратном увеличении концентрации CO2 фотосинтез посева, измеренный при плотности облучения 1600 мкМ/(м2*с), повышался в 2,6 раза. Отмеченный выше линейный характер зависимости фотосинтеза посева от освещенности сохранялся при вариациях дневной/ночной температуры от 25/18 до 37/30 °C. Интересно, что при двукратном повышении концентрации CO2 при выращивании (с 330 до 660 мкМ/моль) нетто-фотосинтез посева увеличился с 60 до 85 мкМ CO2/(м2*с), т. е. примерно на 40%. Расчет данных модели, согласно которой зависимость суточного фотосинтеза посева от концентрации CO2 описывалась прямоугольной гиперболой, давал величину 36%.
Хорошим индикатором адаптации растений риса к углекислотному режиму выращивания является доля белка РБФК от общего содержания в листе водорастворимого белка. Она снизилась на 32% при увеличении концентрации углекислоты при выращивании со 160 до 900 мкМ/моль, что могло быть одной из главных причин резкого снижения (на 66%) активности РБФК в расчете на единицу площади листа (см. табл. 13.5). Интересно, что у сои доля белка РБФК в этих условиях не менялась.
Приведенный выше анализ зависимости ИФ от внутриклеточной концентрации CO2 не указывает на возможность снижения фотосинтетической активности листа по мере повышения концентрации CO2 при выращивании. Видимо, РБФК не в такой мере лимитирует фотосинтез риса при обогащении CO2, в какой это предполагалось.
Дыхание. Интенсивность темнового дыхания на единицу площади посева риса возрастала с увеличением концентрации CO2 при выращивании от 160 до 500 мкМ/моль, а при дальнейшем увеличении до 900 мкМ/моль — постепенно снижалась. ИД в расчете на единицу сухой биомассы в онтогенезе экспоненциально снижалась, и при нормальной и субнормальных концентрациях CO2 была выше, чем при концентрации CO2 500—900 мкМ/моль. ИД снижалась в онтогенезе параллельно уменьшению концентрации азота (r2 = 0,91), т. е. определялась содержанием белка в растении. Кроме того, повышенные концентрации CO2 стимулируют синтез дополнительного количества структурных и неструктурных углеводов в тканях растений, т. е. большая доля сухой массы включалась в небелковые материалы.
Транспирация и эффективность использования воды. Суточное поглощение CO2 и потери воды при транспирации и ЭИВ показаны в таблице 13.7.

Известно, что устьичная проводимость с повышением концентрации CO2 снижается, что может привести к уменьшению потерь воды при транспирации. Однако обогащение воздуха CO2 одновременно может повысить площадь листьев посева, и, соответственно, транспирацию, нивелируя, таким образом, ожидаемую экономию воды. ИЛП увеличился с 7,6 до 10,8, а ЭИВ с 1,2 до 3,9 ммоль СO2/моль H2O при повышении концентрации CO2 от 160 до 900 мкМ/моль. Повышение ЭИВ обусловлено снижением ИТ и одновременным повышением ИФ до концентрации CO2 500 мкМ/моль.
Что касается температуры, то при ее повышении с 28 до 37 °C ЭИВ снижается в результате резкого повышения ИТ при достаточно постоянном значении ИФ в разных температурных условиях. Если бы в качестве показателя эффективности использовали не соотношение интенсивностей фотосинтеза и транспирации, а продуктивность транспирации (биомасса семян, сформированная на единицу израсходованной воды), последняя бы с увеличением температуры резко падала не только из-за повышения потерь воды, но, главным образом, из-за заметного снижения семенной продуктивности.
Таким образом, повышение концентрации CO2 в атмосферном воздухе может благоприятствовать производству риса благодаря активации фотосинтеза, росту урожайности при одновременном снижении транспирационных потерь, а также повышению ЭИВ. Однако в странах с жарким климатом будущее глобальное потепление может привести к заметному снижению урожайности из-за чувствительности цветения и закладки семян к высоким температурам, а также дефицита влаги вследствие повышенной эвапотранспирации.
Пшеница. Фотосинтез прост. При длительном произрастании пшеницы в условиях повышенной концентрации CO2, эффект от начальной активации фотосинтеза (29—98%) и ингибирования дыхания (40—60%) исчезает. Активация фотосинтеза микроценоза пшеницы в атмосфере с повышенным содержанием CO2 при одновременном снижении дыхания в первой половине вегетации (10—13 дней) приводит к увеличению величины нетто-фотосинтеза и стимулирует ростовые процессы, в том числе в листьях. Высокий уровень минерального, в частности азотного, питания позволяет при этом сформировать в 2 раза большую площадь листьев. Такие растения начинают отставать в росте от растений, произрастающих при нормальной концентрации CO2. Одна из причин этого — уменьшение эффективности работы (или скорости регенерации) рибулезобисфосфат-карбоксилазы/оксигеназы (РБФК/о). Теоретически интенсивность карбоксилирования РБФК/о может повышаться на 20—30% при увеличении концентрации CO2 при одновременном торможении оксигеназной активности на 40%. Однако в действительности при длительном воздействии на посевы пшеницы высокой концентрации CO2 наблюдается уменьшение активности или количества РБФК, что объясняется снижением интенсивности синтеза самого белка РБФК/о в результате ограничения доступности азота и недостатка АТФ.
Реакция растений на повышение CO2 в более поздний период вегетации выражена слабее. Высокие температуры, активизируя дыхание растений, способствуют увеличению соотношения дыхания и фотосинтеза. Одновременно из-за увеличения площади листьев возникает конкуренция за свет, и затененные нижние листья активно используют повышенное количество ассимилятов, синтезируемое растениями пшеницы благодаря высокому уровню CO2 в воздухе. В этот период особенно важны устойчивость сорта к затенению и высокий уровень азотного питания, позволяющие сохранять продуктивный стеблестой до уборки урожая.
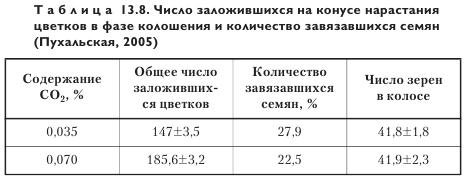
Потенциал продуктивности пшеницы в атмосфере, обогащенной углекислым газом. Уже у ювенильного растения пшеницы, помещенного в атмосферу с высоким содержанием CO2, в конусе нарастания начинает закладываться больше колосков и цветков. Зерновая продуктивность индивидуального растения зависит от гибели части колосков и цветков, заложившихся на VI—VII этапах органогенеза (трубкование-колошение). Чем больше колосков закладывается на конусе нарастания, тем более сильная конкуренция возникает между ними в процессе роста за метаболиты и тем больше колосков гибнет. Высокий уровень CO2 в воздухе не оказывает влияния на эту закономерность (табл. 13.8).
На закладку колосков на конусе нарастания существенное влияние оказывает взаимодействие двух факторов: CO2 — уровень азотного питания. На высоком уровне азотного питания число закладывающихся на конусе нарастания цветков больше. В оптимальных по температурному режиму и водообеспеченности условиях высокая потенциальная продуктивность реализуется в урожае.
Продуктивность растений и азотное питание. У пшеницы существует четкий компенсаторный механизм роста в ценозе, который зависит от концентрации CO2, освещенности и конкурентоспособности растений. У относительно менее требовательных к уровню освещенности сортов, устойчивых к недостатку света, увеличение концентрации CO2 положительно сказывается на росте главного побега, и, по-видимому, из-за этого рост боковых побегов недостаточно обеспечен метаболитами.
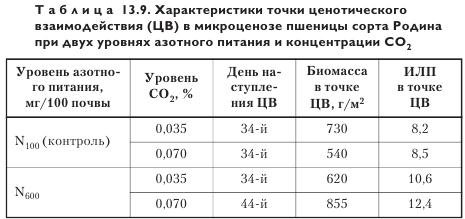
При увеличении концентрации CO2 точка ценотического взаимодействия (ЦВ), отражающая момент начала активной конкуренции за свет, у яровой пшеницы сорта Родина наступает при накоплении меньшей биомассы (665 г/м2) и в более ранней фазе развития, чем при нормальной концентрации CO2 (830 г/м2). Кроме того, повышенная концентрация CO2 способствует удлинению периода жизни некоторых листьев главного побега.
При улучшении азотного питания точка ЦВ наступает при более низких величинах накопленной биомассы, что естественно, поскольку связано с большей величиной ассимиляционной поверхности листьев. Увеличение уровня CO2 на высоком агрофоне обусловливает более позднее начало ценотического взаимодействия (табл. 13.9), т. е. при обогащении CO2 эффективность использования посевом ФАР повышается только при условии достаточного азотного питания.
На низком фоне азотного питания увеличение концентрации CO2 приводит к снижению числа побегов растений пшеницы на единице площади и массы зерна с колосьев главного побега (табл. 13.10).
Повышение уровня азотного питания благодаря улучшению продуктивного побегообразования — необходимое условие увеличения урожая зерна при повышении уровня CO2 в воздухе, поскольку для эффективного использования CO2 необходимо достаточное количество азота в почве. При этом наблюдается повышение не только общей, но и зерновой продуктивности.

Поглощение и вынос элементов питания. Повышенная концентрация CO2 активизирует рост, начиная со второй недели вегетации. Возрастающая конкуренция между растениями за свет и питательные вещества — основной регулирующий рост фактор в ценозе в течение вегетации. К третьему этапу органогенеза (кущение) при содержании CO2 0,07% из-за повышенной ростовой активности проявляется недостаточность азотного питания. Высокая активность основного фермента азотного метаболизма — нитрат -редуктазы — в листьях сохраняется при высоких уровнях CO2 и азотного питания. При низком уровне азота в почве активность нитратредуктазы существенно снижается. Вместе с тем повышенная концентрация CO2 в воздухе не вызывает изменения содержания азота в растении при условии сохранения достаточного для развития корневой системы объема почвы.
С увеличением продуктивности растет и потребность растений в питательных веществах, поскольку при повышении концентрации CO2 увеличивается масса не только зерна, но и вегетативных органов. Накопление азота в растениях пшеницы существенно повышалось как на низком (74%), так и на высоком (45%) уровнях азотного питания. В условиях засухи увеличение накопления азота в растении отмечено только на высоком уровне азотного питания. При созревании пшеницы в условиях повышенного уровня CO2 снижается реутилизация питательных веществ из листьев и содержание белка в зерне. Накопление фосфора растениями увеличивалось при обогащении атмосферы CO2 на 120 и 140% соответственно и на низком, и высоком уровне азотного питания.
Активизация роста и развития корневой системы благодаря стимуляции фотосинтетической деятельности растений при содержании CO2 в воздухе 0,07% приводит к усилению поглощения, а следовательно, и выноса питательных элементов из почвы. Важно, что это происходит без нарушения соотношения макроэлементов.
Устойчивость растений к засухе при повышении концентрации углекислого газа в воздухе. Экспериментально установлено, что в условиях водного дефицита транспорт ассимилятов в листе при обычной концентрации CO2 нарушается раньше, чем в опыте с концентрацией CO2 0,07%. У растений контрольной группы (0,035% CO2) уже на пятый день засухи наблюдалось снижение содержания сахарозы в листьях. К концу засухи, напротив, отмечено резкое возрастание ее содержания в результате нарушения оттока. При обогащении атмосферы CO2 снижение содержания сахарозы наблюдалось только к десятому дню засухи.
В атмосфере, обогащенной CO2, избежать падения водного потенциала листа в условиях нарастающей засухи растению легче, поскольку увеличивается размер корневой системы, что позволяет растению добывать воду из нижележащих слоев почвы. Кроме того, меньшая величина устьичной проводимости ограничивает потерю воды.
В первой половине вегетации (до 36 дней) при условии оптимальной влагообеспеченности суммарное испарение влаги с поверхности микроценоза в атмосфере, содержащей 0,07% CO2, ниже чем в контроле на 40%, а интенсивность транспирации единицы листовой поверхности ниже на 60%, что приводит к повышению ЭИВ. В дальнейшем интенсивный рост листьев на боковых побегах и увеличение площади листьев в опытном варианте приводят к резкому (на 25%) повышению испарения с поверхности посева, что объясняется замедленным старением листьев.
Таким образом, реакция растений на повышение концентрации CO2 при выращивании видоспецифична. Она может заметно измениться при варьировании других атмосферных факторов (концентрация озона, доза УФ-В, содержание поллютантов в атмосфере и др.).