



Влияние ультрафиолетового излучения на физиологические процессы в растении
Ультрафиолетовое излучение (УФ) является важным экологическим фактором, влияющим на растения. Оно входит в состав электромагнитного излучения Солнца и составляет около 7% общей солнечной радиации, достигающей поверхности Земли.
Диапазон УФ-спектра делят на три части: А (Л = 400—320 нм), В (Л = 320—280 нм) и С (Л = 280—180 нм). Излучение с длиной волны менее 295 нм (УФ-С) полностью поглощается озоновым слоем, тогда как УФ-А и УФ-В достигают поверхности Земли. В связи с тем что фотоны УФ имеют энергию, достаточную для ионизации атомов, они поглощаются всеми биологически важными структурами, определяя высокую биологическую активность УФ.
Озоновый слой находится на высоте 10—50 км, максимальная концентрация озона приходится на высоту 20—25 км. Озон (O3) постоянно образуется в верхних слоях атмосферы при комбинации молекулярного и атомарного кислорода. Атомарный кислород возникает вследствие фотодиссоциации O2 коротковолновой УФ-С радиацией (Л < 240 нм). При поглощении УФ молекула O3 разрушается с образованием исходных продуктов. Образование и разрушение озона сбалансировано и является эффективной защитой от УФ. Если учесть колебания мощности озонового слоя, происходящие с циклом 10—12 лет и приводящие к изменению озонового экрана на 10%, можно ожидать наступления периодов весьма значительного уменьшения толщины озонового слоя.
Увеличение поступления фреонов в атмосферу за последние 30—40 лет привело к редукции озонового экрана на 30%. Последствия данных изменений можно представить себе на следующем примере. Так как за пределами атмосферы плотность потока УФ равна 16 Вт/м2, то при полном отсутствии озона в атмосфере мощность поглощенной дозы составляла бы около 1 Гр/ч. При средней дозе LD50 и мощности излучения 30—50 Гр гибель растений наступит через 2—3 сут непрерывного облучения.
УФ-чувствительность экотипов одних и тех же видов растений возрастает по мере смещения места их произрастания из тропических к полярным широтам. УФ-чувствительность растений существенно варьирует не только между видами, но и между различными генотипами одного вида. Относительно резистентными к УФ оказались 12 видов растений (из тестированных к настоящему времени 80), остальные — чувствительными. Определение величины потери урожая полевых культур после УФ-стресса позволило выделить следующие группы растений: устойчивые — райграс, пшеница, овес, кукуруза, подсолнечник; среднеустойчивые — капуста, ячмень, рис; чувствительные — картофель, бобы, горох, вика, клевер, ячмень, огурцы. В настоящее время УФ-чувствительность не рассматривается в качестве селекционного фактора, что может вызвать существенные потери урожая в будущем.
Эффекты, вызываемые УФ, обнаруживаются на всех уровнях организации растений. В основе всех нарушений, наблюдаемых на клеточном уровне, лежат механизмы повреждения биологических молекул. Нарушения обусловлены в основном фотохимическими изменениями пиримидиновых оснований в ДНК, а также фотоинактивацией белков. Последний процесс происходит преимущественно из-за поглощения УФ триптофаном и сопутствующих этому фотохимических реакций. Вследствие нарушения структуры ингибируется ферментативная, регуляторная, транспортная и другие функции белков.
УФ вызывает фотоповреждение белков и фосфолипидов плазматических мембран: окисление липидов мембран по свободнорадикальному механизму с образованием гидропероксидов с последующим их фотохимическим расщеплением и получением стабильных конечных продуктов. Среди ферментов наиболее чувствительной мишенью плазматической мембраны является К+-активируемая АТФаза, инактивация которой может приводить к изменению проницаемости мембран для ионов. Кроме белков, липидов и ДНК в результате поглощения УФ теряют активность витамины, подвергаются деструкции фитогормоны (в первую очередь ИУК), антиоксиданты и другие биологически активные вещества.
Из всего диапазона негативного воздействия УФ на растения главную угрозу представляют мутагенное и летальное действия, в основе которых лежит главным образом повреждение ДНК. При этом ингибируется способность к репликации и транскрипции, возникают нарушения иРНК, ингибируется процесс взаимодействия ее с рибосомами и тРНК, утрачивается трансляционная активность, искажается матричный смысл. При прямом повреждении тРНК подавляется ее акцепторная активность к аминокислотам, изменяется структура антикодона и способность комплексоваться с рибосомами и кодонами иРНК. УФ вызывает разнообразные мутации (структурные мутации хромосом, генные мутации) или нарушения различных физиологических процессов и гибель (как следствие мутаций). Одной из возможных причин мутагенеза при действии УФ является индуцибельная репликация. Известно, что пиримидиновые димеры блокируют синтез ДНК конститутивными ДНК-полимеразами. Однако в условиях индукции повреждений ДНК свойства ДНК-полимераз меняются таким образом, что эти ферменты становятся способными производить неспецифическую застройку матрицы, имеющей в структуре повреждения типа пиримидиновых димеров.
Определен и охарактеризован гиперчувствительный к УФ и ионизирующему излучению мутант арабидопсиса (uvhl), гиперчувствительность у которого обусловлена дефектами в системе репарации ДНК.
Действие УФ на растения можно наблюдать наиболее отчетливо в естественных условиях, например в высокогорье, где интенсивность естественной УФ высока. Здесь растения характеризуются ксероморфной структурой: наличием толстой кутикулы, большого числа слоев паренхимных клеток, увеличением числа клеток, устьиц и хлорофилла на единицу поверхности листа, малыми размерами самих клеток и хлоропластов, а также увеличением количества гран в хлоропластах, тилакоидов в гране, объема тилакоидов в хлоропластах.
С помощью методов генетической инженерии получены растения, которые характеризуются утолщенным кутикулярным слоем, наличием большой опушенности, увеличенным содержанием эпикутикулярных восков, что привело к ослаблению повреждающего влияния на них УФ.
Для защиты растений от УФ важен уровень поглощения последнего в эпидермисе листа. Целлюлоза и гемицеллюлоза, входящие в состав клеточных стенок, проницаемы для УФ, поэтому их экранирующее значение невелико. Вместе с тем лигнифицированные стенки способны поглощать до 50% приходящего УФ. Таким образом, эпидермис функционирует как селективный фильтр солнечной радиации. В среднем эпидермис пропускает около 2% падающего УФ в зонах с высокими уровнями УФ (экватор) и около 5% — в более умеренных широтах. Благодаря дополнительной абсорбции УФ-В в мезофилле и наличию флавоноидов и других поглощающих соединений происходит значительное ослабление потока УФ-В в тканях листа.
Так, флавоноиды ослабляют поток УФ более чем на 90%. Значительное количество флавоноидов содержится в вакуолях клеток эпидермиса и мезофилла, обнаружены они и в хлоропластах некоторых видов растений, где также выполняют роль фильтра УФ.
УФ влияет на устьичную проводимость водяных паров и углекислого газа в лист и соответственно — на фотосинтез, а также на множество других физиологических процессов — от темнового дыхания и транспорта минеральных солей до угнетения роста и развития растений, формирования урожая.
Фотосинтез. УФ прямо ингибирует фотосинтез: нарушает функционирование ФС II, снижает активность ФС I (хотя и в меньшей степени, чем ФС II), разрушает пластохиноны, дезинтегрирует мембранные структуры, ингибирует электронный транспорт, связанный с ФС II, ингибирует систему разложения воды и подавляет реакции Хилла, меняет выход флуоресценции хлорофилла а в изолированных хлоропластах и целых листьях.
Большинство названных эффектов объясняются разрушением структуры мембран УФ и инактивацией компонентов, необходимых для световых и темновых реакций фотосинтеза. Высокий уровень УФ в диапазоне 280—320 нм вызывал затруднения для проникновения CO2 в мезофильные клетки, ограничивая фотосинтетический процесс, что было показано, в частности, на растениях ячменя в условиях высокогорного Памира. Прослеживается связь между снижением интенсивности фотосинтеза и уменьшением открытости устьиц, что приводит к увеличению сопротивления поступления CO2 в лист растения. С3-растения в целом более чувствительны к воздействию УФ, чем С4-растения, что связано с сохранением активности реакции Хилла у последних.
Снижение активности первичных процессов во многом обусловлено структурными изменениями фотосинтетического аппарата, определяемыми в том числе и длительностью экспозиции. В опытах с 30- и 60-минутным облучением ультрафиолетом растений гороха наблюдалось увеличение суммарного хлорофилла на 10—15% через 2—3 сут. После облучения в течение 2 и 2,5 ч имело место снижение содержания хлорофилла на 16 и 10% соответственно. Длительное облучение ультрафиолетом приводило к снижению отношения а/b хлорофиллов в прилистниках гороха, причем это изменение в большей степени было обусловлено уменьшением концентрации хлорофилла а, особенно при низкой интенсивности света. Интересно, что подавление синтеза хлорофилла а наблюдалось на ранних этапах развития растений, а хлорофилла b — на поздних. Однако только фотодеструкцией хлорофиллов нельзя объяснить снижение фотосинтетической активности, поскольку степень подавления реакции Хилла заметно сильнее снижения концентрации хлорофилла.
Изменение содержания хлорофилла у растений, выращенных при высокой интенсивности облучения, совпадало с увеличением накопления сухого вещества прилистниками (на 17% по сравнению с контролем), однако увеличение площади при этом было незначительным. Наблюдаемые изменения являются приспособительной реакцией фотосинтетического аппарата, направленной на уменьшение повреждения клеток мезофилла. Стимулирующий эффект сменялся ингибированием ростовых процессов при достижении продолжительности облучения 2,5 ч.
Устьица. Хотя эффект УФ-В на восстановление углерода прямо не опосредован диффузией CO2 в листе, дополнительная доза облучения может косвенно лимитировать фотосинтез путем индукции закрывания устьиц в результате ингибирования накопления K+ в замыкающих клетках. Устьичная проводимость заметно снижалась уже через 2 нед. воздействия относительно низкой дозой УФ-В на растения фасоли, сои и огурца. Огурец отличается особой чувствительностью, теряя все устьичные функции уже через 9 дней облучения. Однако реакция устьиц в значительной мере определяется условиями среды. Так, при прямом действии УФ-В устьица закрываются, открываясь вновь при последующем воздействии белым светом. Кроме того, если в условиях оптимальной влагообеспеченности УФ-В индуцирует закрывание устьиц, при дефиците влаги этого не наблюдается. Косвенное влияние УФ-В на устьица может также заключаться в снижении их числа на единицу площади листа на фоне уменьшения площади листьев и увеличения их толщины. Исследования показывают, что прямое ингибирование фотосинтеза УФ-В не связано с устьичным лимитированием. В опытах с дискриминацией изотопов углерода показана более высокая внутриклеточная концентрация CO2 при облучении УФ-В, что также указывает на прямое ингибирование фотосинтеза. Таким образом, в отсутствие прямых изменений устьичной проводимости, УФ-В индуцирует хроническое снижение фотосинтетической способности, что может происходить в разных звеньях цикла восстановления углерода.
Фотосистемы I и II. Хотя ФС I может ингибироваться при освещении хлоропластов интенсивной УФ-С, последняя не присутствует в естественной среде. Что касается УФ-В, то в большинстве случаев ее влияние на ФС I гораздо слабее, чем на ФС II. Так, на выделенных хлоропластах гороха показано, что УФ-В, полностью ингибировавшая ФС II, снижает скорость циклического фотофосфорилирования через ФС I только на 35%. Это дает основание считать, что ингибирование фотосинтеза связано прежде всего с ФС II. Интересно, что в параллельных опытах на горохе показано, что содержание ФС I и цитохрома f на единицу площади листа снизилось на 58%, а активность ФС II — на 80% при неизменности первых двух параметров в расчете на единицу хлорофилла.
Реакционный центр (РЦ) ФС II содержит две формы мембранных белков с молекулярными массами 32 и 34 кДа, известных соответственно как Д1 и Д2. Хлорофилл а, феофитин, пластохинон связаны с этими белками. Сам реакционный центр P680 повреждается повышенной УФ-В как в отношении окисления воды, так и восстановления пластохинона. Д1-белок комплекса РЦ особенно чувствителен к УФ-В, вызывающего быструю его деградацию.
Акцепторная, или восстанавливающая, часть белков Д1 и Д2 также может быть модифицирована УФ-В, которая изменяет также кинетику флуоресценции, причем быстрые ее компоненты ускоряются, а медленные тормозятся. Предполагается также, что УФ-В вызывает прямое повреждение молекул пластохинона, которые могут выступать в качестве первичных сенсоров действия УФ-В.
Активность и содержание РБФК/о. В целом ряде исследований показана обратная корреляция между интенсивностью облучения УФ-В и активностью РБФК. Активность ФЕПК у кукурузы (СД-растения) тоже снижалась, но при очень высоком уровне УФ-В. Косвенно этот вывод подтверждается при анализе A/сi кривой фотосинтеза, угол наклона линейной части которой при дополнительном УФ уменьшается.
УФ-В снижает концентрацию белка и РБФК/о в листе. Так, УФ-В, соответствовавшее 36%-му истощению стратосферного озона, снижало как активность, так и количество РБФК. В одном из опытов максимальная активность РБФК снизилась на 71% после воздействия УФ-В, имитирующего почти 3-кратное превышение над естественным фоном в течение 3 сут. Интересно, что уровень транскриптов мРНК, кодирующих малую (rbcS) и большую (rbcL) субъединицы РБФК, резко снизился в течение нескольких часов воздействия УФ-В, причем для rbcS на 20% в сравнении с контролем. Однако влияние УФ-В на транскрипты мРНК обратимо. Вышесказанное, как и данные о влиянии УФ-В на ген psbA, кодирующий хлоропластный белок Д1, указывает на возможную быструю регуляцию экспрессии генов при действии ультрафиолетового излучения.
Водный режим. Нарушение функционирования устьиц в результате воздействия УФ-В повышенной интенсивности приводит к увеличению потери воды за счет транспирации и вызывает водный стресс и увядание растений, подавляющих фотосинтез. Комбинирование воздействия УФ и умеренной засухи ингибирует фотосинтез у молодых растений в большей степени, чем воздействие каждого из рассматриваемых факторов в отдельности, т. е. наблюдается синергический эффект. Взрослые растения менее чувствительны к УФ-В, чем к засухе.
Важную роль в поддержании водного баланса играют поверхностные (эпикутикулярные) воски, являющиеся барьером для диффузии воды в листьях растений. Их удаление снижает отражающую способность листьев приблизительно в 2 раза с соответствующим увеличением поглощения света. УФ-В вызывает сдвиг распределения основных компонент поверхностных восков в сторону более коротких цепей, причем этот эффект обусловлен прямым действием на их биосинтез.
Минеральное питание. Отмечена более слабая чувствительность к УФ-В растений, произрастающих в условиях дефицита фосфора. УФ-В не оказывало какого-либо влияния на рост и фотосинтез фосфордефицитных растений. В случае же избытка азота и влаги мезоструктура листьев характеризуется меньшей толщиной эпидермиса и кутикулы, вследствие чего следует ожидать повышения чувствительности растений к УФ-В.
Очень важную, хотя и опосредованную, роль в действии УФ на растения могут играть почвенные микроорганизмы, влияющие на плодородие почв. То обстоятельство, что почва интенсивно поглощает УФ, не исключает его влияния на почвенные микроорганизмы, поскольку большинство из них и практически все фототрофные организмы обитают в верхних слоях почвы. Известно, что УФ-В способно инактивировать ключевой фермент ассимиляции азота — нитрогеназу. Кроме того, УФ повышенной интенсивности, вероятно, вызовет вытеснение и отмирание чувствительных и размножение устойчивых микроорганизмов, часть из которых могут оказаться патогенными.
Рост и развитие. Чувствительность растений к ауксину в результате воздействия повышенной дозой УФ меняется. Ауксин, как известно, контролирует апикальное доминирование и характер листорасположения, что в свою очередь влияет на поглощение растением света. Изменения высоты растений и длины междоузлий могут быть обусловлены индуцированным УФ-В фотоокислением ИУК. Повышенные дозы УФ снижают концентрацию ауксина, в частности, в гипокотилях проростков подсолнечника.
Изменения в концентрации ИУК в результате действия УФ вызывают целый ряд морфологических изменений. Уменьшение длины междоузлий и соответственно низкорослость, в частности проростков, представляет собой типичный пример воздействия УФ, особенно заметный при сочетании высокого уровня УФ и низкой ФАР. Однако даже при уменьшенных дозах УФ-В, имитирующих снижение концентрации озона на 20% на 40° северной широты, у многих полевых культур отмечено сокращение длины междоузлий и листовых пластинок при одновременном повышении массы листьев и стеблей. У чувствительных сортов риса, напротив, в этих условиях снижалось число продуктивных побегов. Изменение доли отдельных органов в биомассе целого растения является типичной реакцией двудольных растений, у которых больше ассимилятов транспортируется в листья и меньше — в стебли и корни. У однодольных, в частности риса, чувствительные сорта, снижая абсолютную массу практически всех органов, не обнаруживают изменений в соотношении масс органов.
Морфологические изменения, индуцированные УФ-В, включая торможение роста одних частей растений и стимулирование других, могут происходить без какого-либо видимого ингибирования фотосинтеза на уровне отдельного листа. Однако изменения этих параметров могут повлиять на поглощение света и привести к физиологическим изменениям на уровне целого растения.
Кроме снижения площади листьев, дополнительная УФ-В у чувствительных культур зачастую на 60—70% повышает толщину листьев и соответственно УПП. Этот эффект достигается за счет увеличения как числа, так и размера клеток палисадной и губчатой паренхимы. Одновременно изменяются свойства поверхности листьев, увеличивая рассеивание и отражение света и соответственно снижая количество световой энергии, доступной для фотосинтеза. Показано, что усиленное воздействие УФ повышает содержание эпикутикулярных восков у полевых культур до 25% при заметном изменении их состава. Кроме влияния на поглощение и рассеивание света, это может уменьшить транспирацию, изменив реакцию фотосинтеза на температуру и влажность.
Одним из типов реакции растений на облучение УФ является фотоморфогенез. Так, УФ-В ингибирует рост гипокотиля мутантов огурца, дефектных по фитохрому. Одним из механизмов участия УФ в фотоморфогенезе является изменение концентрации фитогормонов вследствие фотохимических реакций. В частности, наблюдалось ингибирование растяжения эпидермальных клеток в проростках подсолнечника в результате фотоокисления ИУК.
Замедление роста может быть обусловлено также увеличением содержания в растениях ABК, что вызывает относительно синхронное системное ингибирование функций роста и фотосинтеза, включающее подавление системы репликации, деления и структурной дифференциации клеток и хлоропластов. Для растений, устойчивых как к засухе и газообразным загрязнителям, так и к воздействию УФ, характерны высокий исходный уровень АБК и быстрое увеличение его содержания в результате этих воздействий. Аналогичные закономерности установлены и для другого стрессового гормона — этилена. На фоне действия УФ ослабляется эффект ауксинов и гиббереллинов, причем последние снимают негативное действие УФ.
Торможение роста растений коррелирует с подавлением формирования листовой поверхности. УФ-В вызывает повреждение корневой системы, о чем свидетельствует значительная степень депрессии ее роста. Под микроскопом кончики корней облученных растений выглядят поврежденными: часть корневых волосков лопнула, движения цитоплазмы в корневых волосках нет.
Вместе с тем малая доза УФ (2,1 кДж/м2) продолжительностью 30 мин увеличивает высоту побегов и размеры листьев у опытных растений. Кроме того, увеличиваются объем корневой системы, общая и рабочая адсорбирующие поверхности на фоне снижения содержания растворимых сахаров. Большая доза УФ (14,5 кДж/м2) продолжительностью 30 мин тормозит линейный рост растений пшеницы и бобов, уменьшает размеры листьев, снижает у пшеницы общую адсорбирующую поверхность корней на 29%, рабочую — на 77%, а у бобов на 42 и 47% соответственно. Однако при этом у опытных растений пшеницы почти в 20 раз увеличивается содержание растворимых сахаров; уровень общего азота возрастает на 23% у пшеницы и на 85% у бобов. Таким образом, в зависимости от дозы УФ ответ растения на морфологическом и метаболическом уровнях может иметь как стимулирующий, так и ингибирующий характер.
Формирование урожая и его качество. Цветение и опыление. Сведения о влиянии УФ на цветение и опыление немногочисленны и не создают впечатления возможности радикальных изменений этих функций. Стенки пыльника поглощают около 98% падающего на него излучения. Во время опыления пыльца также хорошо защищена, поскольку ее стенка содержит УФ-поглощающие компоненты. Однако после переноса пыльцы на рыльце пестика пыльцевая трубка может стать чувствительной к УФ-В. Этот эффект более вероятен у растений с бинуклеарным типом пыльцы и с большим временем прорастания по сравнению с тринуклеарным.
В экспериментах с 10 видами растений получены косвенные доказательства подавления прорастания пыльцы в результате воздействия УФ. Даже 1—3-часовое воздействие УФ-В умеренной интенсивности подавляет прорастание пыльцы некоторых видов на 33—65%; у 3/4 видов эффективными оказываются интенсивности облучения, характерные для высокогорья или тропиков. Яйцеклетки хорошо скрыты в завязи и достаточно защищены от естественного УФ.
Согласно другим сведениям, пыльца растений до момента раскрытия пыльников полностью защищена от УФ венчиком цветка и стенками пыльника. Вероятность попадания пыльцы под действие УФ в момент опыления у самоопыляющихся растений наименьшая.
У растений, выращиваемых в условиях памирского высокогорья, отмечалось увеличение количества цветочных стрелок, уменьшение интенсивности цветения однолетних растений, превращение кустистых форм в розеточные. В полевых экспериментах с 7 конкурирующими парами культурных растений и сопутствующих им сорняков было обнаружено, что увеличение интенсивности УФ-В повышает конкурентоспособность пшеницы по отношению к сорнякам. Подобный результат в основном обусловлен морфологическими изменениями. В условиях воздействия УФ-В повышенной интенсивности и при одинаковом фотосинтезе высота стебля и длина листьев у дикого овса оказываются меньше, чем у пшеницы.
Урожай и его качество. Анализ ростовых реакций ячменя, проведенный в фазах выхода в трубку и молочной спелости, показал, что растения ячменя к концу вегетации частично адаптируются к действию УФ повышенного уровня. Наиболее чувствительным показателем является ЧПФ. Высокая доза УФ приводит к перераспределению сухой массы в пользу листьев и уменьшению ее поступления в стебель, что может быть причиной полегания и потери урожая (табл. 13.1).

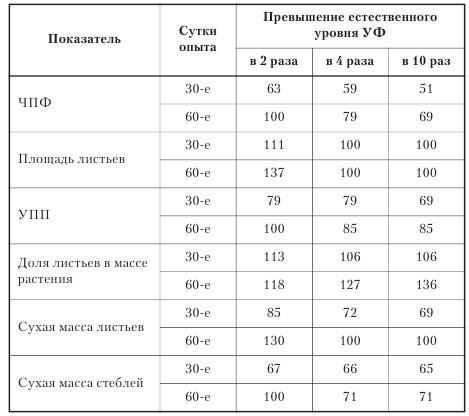
Потери урожая при 2-кратном увеличении естественного уровня УФ составили 8,5%, при 4- и 10-кратном соответственно 25,5 и 38,3% (табл. 13.2).
Основными слагаемыми уменьшения продуктивности являются снижение крупности, а также числа семян, особенно с боковых побегов.


Интересно, что при истощении стратосферного озона на 16, 23 и 32% и соответственно повышении УФ потери урожая риса составили 24, 24 и 48%, а пшеницы — 2, 15 и 13% соответственно. Таким образом, в указанных условиях пшеница оказалась более устойчивой, чем рис. При имитации 32%-го истощения, урожайность сортов сои составила в среднем 97% от контроля при вариациях от 91 до 109%. При имитации менее чем 10%-го истощения озона, урожайность картофеля, шпината, фасоли, редиса, ячменя и капусты не только не убывала, но в ряде случаев даже увеличивалась. В условиях 40%-го истощения урожай картофеля снижался до 41%, шпината (листья) — до 66%, капусты (листья) — до 49% и фасоли (бобы) — до 75%.
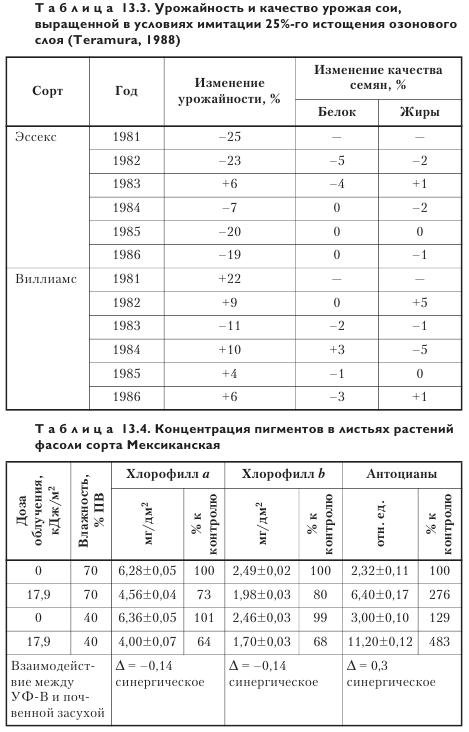
УФ-В повышенной интенсивности, достигаемое путем снижения концентрации озона, может вызвать уменьшение содержания протеина и жиров в бобах, тем самым снижая качество урожая. Этот эффект усиливается в результате накопления в большинстве растений флавоноидов, негативно влияющих на здоровье человека (табл. 13.3).
Зависимость реакции растений на УФ от сочетания факторов среды. Воздействие УФ на фоне сочетания действия разнообразных стрессоров оказывает неоднозначное влияние на физиологические функции растений. В одном из опытов определялось влияние температуры и влажности почвы на чувствительность фасоли к УФ (табл. 13.4).
Обнаружен синергический негативный эффект при действии УФ-стресса и пониженной температуры (12 °C) или УФ-стресса и низкой влажности почвы (40% ПВ). В условиях пониженной температуры (12 °C) содержание антоцианов у облученных растений было почти в 2 раза выше, чем у необлученных, а при оптимальной температуре вегетации (20 °C) — лишь на 17% выше, чем у последних. Недостаточное водоснабжение (40% ПВ) в период, следующий за УФ-стрессом, усиливает его неблагоприятное влияние, способствуя разрушению хлорофиллов и каротиноидов и накоплению антоцианов. В условиях оптимального водоснабжения (70% ПВ) количество антоцианов после действия УФ-стресса возрастало в 2,8 раза, а при недостаточном водоснабжении (40% ПВ) — в 3,7 раза. Само по себе недостаточное водоснабжение увеличивает содержание антоцианов в 1,3 раза. Показано, что УФ повышенного уровня изменяет водный режим растений, нарушая их водный баланс, а признаки, характерные для водного стресса (потеря тургора, обезвоживание), присущи и облученным УФ растениям. Недостаток воды в почве еще более усиливает водный дефицит растений, подвергнутых действию УФ-стресса.
Хотя УФ-В и засуха — факторы, действующие в естественных условиях совместно, устойчивость к ним не всегда сочетается в пределах одного генотипа. Из проанализированных восьми сортов пшеницы лишь два (Саратовская 29 и Альбидум 28) обладали как высокой засухоустойчивостью, так и устойчивостью к действию повышенного УФ-В, в связи с чем они могут служить донорами при селекции на повышенную устойчивость к этим факторам.
Эффект взаимодействия УФ-В и высокой температуры в период вегетации для пшеницы (b = -0,26) и ячменя (b = -0,75) является синергическим: при высокой температуре (28 °C) в июле-августе продуктивность растений пшеницы и ячменя под действием УФ снижалась соответственно на 25 и 54%, тогда как при среднесуточной температуре 18 °C у пшеницы она не изменялась, а у ячменя была меньше на 12%.
Механизмы защиты растений от ультрафиолетового излучения. Для снижения негативного воздействия УФ-В растения выработали ряд защитных стратегий. Так, изменения в анатомии листьев (в частности, толщины и пигментации) защищают фото-синтетический аппарат от избытка УФ-В. Однако эти механизмы обычно сокращают количество поглощаемой листом ФАР и, соответственно, снижают фотосинтез. Остановимся на механизмах репарации, которые уменьшают вызванные ультрафиолетом повреждения без заметного снижения фотосинтетической активности.
Синтез флавоноидов. Длительное время считалось, что флавоноиды, принадлежащие к классу водорастворимых фенольных производных, связаны с защитой от УФ-В. Флавоноиды поглощают исключительно в зоне УФ-В с максимумом поглощения около Л = 300 нм. Исследования на мутантах арабидопсиса показали, что изогенные линии с пониженным содержанием флавоноидов и фенольных компонент были исключительно восприимчивы к действию УФ-В по сравнению с исходными генотипами. Флавоноиды присутствуют в листьях, пыльце, стеблях, но прежде всего в вакуолях и клеточных стенках. Эпидермис листьев создает первый защитный барьер против УФ-В, при этом концентрация флавоноидов в эпидермисе достаточно высокая (до 10 ммоль).
Индукция биосинтеза флавоноидов при повышении уровня УФ-В может регулироваться на генном уровне, поскольку искомый фактор увеличивает концентрацию ключевых ферментов биосинтеза флавоноидов. Имеющиеся данные указывают на возможную роль флавина в качестве фоторецептора. Однако простое увеличение концентрации флавоноидов в листе полностью не исключает отрицательный эффект УФ-В. Так, фотосинтетический аппарат некоторых видов растений, произрастающих в тропических регионах на большой высоте над уровнем моря, достаточно устойчив к повышенному уровню ультрафиолетовой радиации. Однако эта устойчивость далеко не всегда связана с повышенной концентрацией флавоноидов. Поэтому считается, что накопление флавоноидов может вносить определенный вклад в устойчивость, однако простые причинно-следственные связи при этом не всегда могут существовать.
Полиамины. Накопление полиаминов иногда связывают с целым рядом биотических и абиотических стрессов, особенно водным. Однако специфические типы полиаминов, такие, как спермидин, спермин и путресцин, могут снижать ПОЛ, что стимулируется у растений, подвергнутых повышенным дозам УФ-В. Соответственно, полиамины могут быть фактором, вносящим вклад в целостность процессов фотосинтеза при высокой дозе УФ-В.
Следует отметить, что адаптация к низким и умеренным дозам УФ-В проходила на фоне стабильного уровня спермидина и спермина и увеличения содержания их предшественника — путресцина (рис. 13.1). Видимо, накопление путресцина позволяет поддерживать пул спермидина и спермина при адаптации, находящейся под строгим гомеостатическим контролем. Снижение содержания путресцина после высокой и летальной доз УФ-В сопровождалось падением уровня спермидина и спермина, которые интенсивно расходуются в стрессовых состояниях, участвуя в стабилизации биологически важных макромолекул.
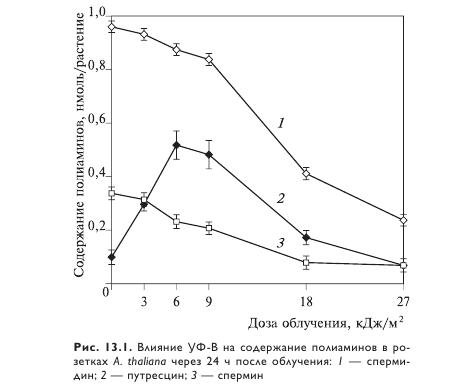
Предполагают, что в растениях существуют по крайней мере два пути трансдукции УФ-В-сигнала, в которых участвуют фитогормоны. Первый, индуцируемый низкими дозами УФ-В, позволяет растениям адаптироваться к радиации и сопровождается транзиторным усилением образования этилена, продолжительным накоплением ABК, увеличением синтеза путресцина и поддержанием стабильно высокого уровня спермидина и спермина. Второй, запускаемый высокими дозами, вызывает продолжительный подъем образования этилена, снижение накопления ABК, резкое падение содержания полиаминов и гибель растений.
Кислородные радикалы и метаболиты. Основные компоненты фотосинтетического аппарата, в частности липиды и белки, восприимчивы к повреждениям различными формами кислорода и свободными радикалами. УФ-В, в свою очередь, может увеличить их концентрацию. Поэтому неудивительно, что в растениях имеется целый ряд систем, защищающих от этих радикалов. Подобные системы включают СОД, а-токоферол (витамин Е), перекись водорода, аскорбат, b-каротин и глутатион. Интересно, что флавоноиды в дополнение к тому, что они ослабляют негативное влияние УФ-В, могут также играть ключевую роль в качестве антиоксидантов, препятствующих стимулируемым кислородом окислительно-восстановительным реакциям в хлоропласте.