



Токсическое действие алюминия на растения
Воздействие на растения. Алюминий достаточно быстро действует на все клеточные функции (рис. 12.3), поэтому сложно отделить его первоначальный эффект на клетки растений от следствия, определив причину токсичности. Поглощение алюминия отчлененными корнями сельскохозяйственных культур (пшеницы, фасоли, капусты, лука) демонстрирует двухфазную кинетику. Быстрая нелинейная неметаболическая фаза поглощения протекает в течение первых 20—30 мин. Предполагается, что эта фаза обусловлена накоплением алюминия в апопласте клеток. Следующая за ней метаболическая линейная фаза связана с поглощением алюминия симпластом. В течение первых трех часов воздействия происходило накопление алюминия в клеточной стенке, где в этот период времени подавляющая часть алюминия и оставалась. Поступление алюминия в протопласт происходило незамедлительно, но составляло лишь 0,05% от общего накопления алюминия. Поступление в вакуоль происходило с 30-минутной лаг-фазой.
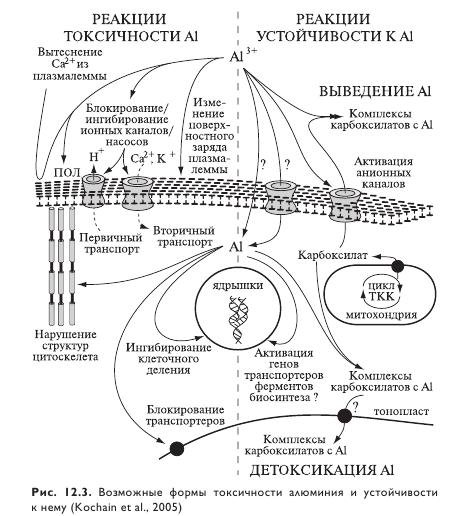
У молодых проростков наиболее чувствительным к алюминию является корневой апекс, точнее дистальная часть проводящей зоны апекса. Воздействие алюминия на любую другую часть корня не влияет на его рост. Кончик корня аккумулирует сравнительно больше алюминия, чем другие его части. Небольшое количество алюминия обнаруживается в цитозоле уже после нескольких минут экспозиции. Хотя большая часть алюминия аккумулируется в апопласте, небольшая фракция все-таки поступает в симпласт, взаимодействуя там с мишенями.
Интересно, что ингибирование роста клеток корней растяжением происходит очень стремительно (в пределах минуты), в то время как подавление клеточного деления проявляется лишь через 6—24 ч. Клетки в дистальной части зоны проведения корневого апекса кукурузы, которые недавно поделились, но еще не начали растягиваться, являются наиболее активными в поглощении алюминия и наиболее чувствительными к его токсическому воздействию. Из-за высокого содержания пектина стенки клеток этой зоны аккумулируют алюминий быстрее, чем стенки более старых клеток. Связываясь клеточными стенками корня, алюминий вытесняет кальций из пектиновой фракции, что в свою очередь подавляет растяжение клеточной стенки. Кроме того, алюминий может быстро ингибировать транспорт ауксинов в клетки зоны проведения у чувствительного к алюминию генотипа кукурузы, что может быть причиной подавления роста корней.
Растения некоторых видов отвечают на действие алюминия повышенным синтезом гемицеллюлозы, целлюлозы и пектина. Однако, удерживая алюминий в апопласте, эти углеводы могут в дальнейшем препятствовать росту клеток растяжением. Индуцированное алюминием ингибирование поглощения K+ путем блокирования калиевых каналов также может препятствовать тургорзависимой элонгации клеток. И, наконец, растущие клетки корня в ответ на воздействие алюминия аккумулируют каллозу на плазма-лемме. Поэтому ее накопление в апопласте используется в качестве раннего симптома токсического действия алюминия. Синтез каллозы сильно зависит от присутствия Ca2+, и его вытеснение с поверхности мембраны алюминием может повышать пул Ca2+ в апопластном пространстве, стимулируя синтез каллозы. Поэтому не исключено, что ингибирование роста корней является отчасти следствием использования сахаров в качестве субстрата для образования каллозы вместо целлюлозы. В свою очередь, каллоза, откладываясь в плазмодесмах, может заблокировать межклеточный транспорт.
Помимо быстрой аккумуляции в клеточных стенках и апопласте корней, алюминий у неустойчивых видов растений накапливается в плазматической мембране и в симпласте, влияя на многие этапы роста корней. Именно плазматическая мембрана была предложена в качестве первичной мишени токсичного действия алюминия. Прочно связываясь с плазмалеммой, алюминий вызывает ее деполяризацию, что в свою очередь запускает последующий каскад событий. Это хорошо показано на примере клеток наружного корового слоя зоны проведения корней чувствительных к действию алюминия генотипов кукурузы. Что касается поступления алюминия непосредственно в цитоплазму, это, видимо, происходит путем трансмембранного переноса с участием кальциевых каналов. После 30-минутной экспозиции у чувствительного генотипа алюминий накапливается в симпласте, а также в ядрах клеток корневого апекса. Уже через 1 ч в этих клетках дезинтегрировались микротрубочки цитоскелета, и каллоза накапливалась в клеточных стенках. На суспензионной культуре клеток сои алюминий быстро увеличивал жесткость трансвакуолярной актиновой сети. Поэтому не исключено, что именно цитоскелет является первичной мишенью алюминиевой токсичности у растений. Высказано предположение, что фосфорилированные сайты на миозине или других актинсвязывающих белках могут связывать и алюминий, предотвращая доступ последнего к фосфатазам и стабилизируя актиновую сеть. Установлено, что алюминиевая токсичность у пшеницы вызывает повышенную экспрессию гена, кодирующего актинсвязывающий белок, участвующий в поддержании функции цитоскелета.
Алюминий может также нарушить процесс сборки и разборки микротрубочек путем ингибирования гидролиза инозинтрифосфата и снижения чувствительности к регуляторным сигналам от Ca2+. Исчезновение микротрубочек наблюдалось иногда в клетках зоны растяжения корней пшеницы при 3-часовом действии алюминия в концентрации 50 мкМ. В наружных клетках коры дистальной транзитной зоны корней чувствительной к алюминию кукурузы микротрубочки исчезали через 1 ч после начала действия алюминия в концентрации 30 мкМ. При этом микрофиламенты оказались менее чувствительными к алюминию.
Кроме того, алюминий быстро увеличивал концентрацию свободного кальция в цитозоле — Caцит2+. Нарушения в кальциевом гомеостазе и ингибирование алюминием активности фосфолипазы С возможно влияют на структуру и функции цитоскелета. Ингибирование фосфолипазы С может также нарушать биохимический путь трансдукции сигналов.
Первичные биохимические нарушения в результате действия алюминия могут быть обусловлены формированием комплекса алюминия с кальмодулином — Са2+-зависимым регуляторным белком. Кальций, являясь вторичным мессенджером, запускает в клетках каскад процессов для противодействия токсичности алюминия. При концентрации 50 мкМ алюминия в корнях пшеницы отмечено осцилирующее увеличение концентрации Ca2+ в цитозоле. Аналогичное отмечено и в клетках зоны растяжения корня у чувствительного к алюминию сорта ячменя. Поскольку известно, что алюминий блокирует кальциевые каналы, позволяющие кальцию поступать в цитоплазму, видимо, Ca2+ высвобождается из внутриклеточных запасных пулов. Следует особо подчеркнуть, что повышение концентрации цитозольного Ca2+ всегда предшествует алюминийзависимому образованию каллозы — быстро проявляющемуся индикатору алюминиевой токсичности.
Токсичное действие алюминия после остановки растяжения клеток связано с изменениями совокупности физиологических процессов и биохимических путей. В силу высокой реакционной способности алюминий обладает сильным сродством к протеинам, неорганическим фосфатам, нуклеотидам, РНК, ДНК, карбоновым кислотам, фосфолипидам, флавоноидам и антоцианам. Симптом токсичности связан с образованием комплексов алюминия с карбоксильными группами пектиновых веществ клеток корня или с переключением синтеза целлюлозы на аккумуляцию каллозы, а также ингибированием митоза в корневом апексе с последующей остановкой синтеза ДНК, хромосомными абберациями, появлением мостов в анафазе, слипанием хромосом и программируемой смертью клеток в апексе корня, запускаемой АФК. Образование АФК и сопряженные с ними реакции ПОЛ на более поздних этапах действия алюминия, как и окислительный стресс, возможны благодаря тому, что активность митохондрий под действием алюминия подавляется, в результате чего ингибируется дыхание и синтез АТФ.
Таким образом, в силу сложности множества процессов, вовлеченных в ингибирование роста корней, конкретные мишени токсичного воздействия алюминия во всей этой сложной цепочке событий все еще остаются неясными. Очевидно лишь то, что наиболее значимое место среди них занимают клеточная стенка, плазматическая мембрана, пути трансдукции сигналов, цитоскелет клеток корня, ДНК/ядро клетки. Вместе с тем понимание механизмов токсичности алюминия и устойчивости к нему растений является важным при разработке соответствующих тестов для отбора толерантной зародышевой плазмы и создания сортов растений с повышенной устойчивостью. Рассмотрим далее, что же происходит в таких приграничных структурах, как клеточная стенка и плазмалемма.
Клеточная стенка. Значительная часть алюминия в корнях взаимодействует со связывающими сайтами апопласта, преимущественно в клеточных стенках периферии корня. Суммарный отрицательный заряд клеточной стенки определяет ее катионообменную способность и, следовательно, возможность взаимодействия алюминия с клеточной стенкой. Среди множества компонентов клеточной стенки именно пектины были предложены в качестве наиболее важного сайта указанного взаимодействия, при котором происходит вытеснение других катионов, в частности Ca2+, необходимых для прочности клеточной стенки. Быстрое связывание алюминия может изменить структурные и механические параметры клеточной стенки, делая ее более жесткой и снижая механическую растяжимость, необходимую для нормального роста клеток.
Алюминий наряду с кальцием вытесняет и магний из стенок клеток коровой паренхимы. Пектин выделяется в форме эфиров из симпласта в апопласт, где происходит деметилирование с помощью фермента пектинметилэстеразы. В результате высвобождаются карбоксильные группы для последующего связывания с алюминием. Трансгенный картофель, сверхэкспрессирующий данный фермент, более чувствителен к алюминию, что определяется более сильным подавлением роста корней по сравнению с немодифицированными растениями. Эти данные могут свидетельствовать о том, что число мест связывания алюминия в апопласте положительно коррелирует с чувствительностью к нему.
Кроме внешнего связывания с компонентами клеточной стенки, алюминий может вмешиваться в механизмы синтеза или отложения полисахаридов в клеточной стенке. Воздействие 10 мкМ алюминия в течение 6 ч уменьшало растяжимость клеточной стенки корней проростков пшеницы. При этом модифицируются компоненты клеточной стенки: увеличивается молекулярная масса гемицеллюлозных полисахаридов и уменьшается эластичность клеточных стенок, что соответственно ограничивает их растяжимость. Содержание уридиндифосфатглюкозы (УДФГ) — субстрата для синтеза целлюлозы в корнях кукурузы после 20-часовой экспозиции при 0,1 мМ алюминия снижалось на 65%, что тоже могло ингибировать образование полисахаридов клеточной стенки.
Плазматическая мембрана. Фосфолипиды в составе мембраны содержат фосфат, и алюминий может присоединиться к этой отрицательно заряженной группе. Фосфатидилхолин — наиболее часто встречаемый в плазмалемме растений фосфолипид, и Al3+ обнаруживает in vitro более чем 560-кратное преимущество по сродству с поверхностью этого фосфолипида по сравнению с Ca2+. Липиды с общим отрицательным зарядом головок, в частности фосфатидилинозитол, обладают большим сродством к алюминию, чем фосфатидилхолины, головки которых нейтральны. В этой связи следует подчеркнуть, что экспрессия кДНК — Ta PSS1, кодирующей фосфатидилсеринсинтазу (PSS), усиливалась в ответ на повышение концентрации алюминия в корнях.
Связывание Al3+ с наружной поверхностью фосфолипидов снижает поверхностный отрицательный заряд мембран. Обнаружена тесная корреляция между относительным ингибированием роста корня у чувствительного к алюминию сорта пшеницы и активностью алюминия на поверхности мембраны (но не в растворе). Аналогичная тенденция обнаружена на корнях тыквы. Путем измерения поглощения радиоизотопов корнями ячменя показано, что в присутствии алюминия поглощение катионов (К+, NH4+ и Ca2+) уменьшалось, а анионов (NO3, HPO2-) увеличивалось. He исключено, что присоединение алюминия к наружной поверхности плазмалеммы формирует поле положительных зарядов, что тормозит движение катионов к поверхности мембран и, напротив, стимулирует движение анионов. Вместе с тем Mg2+ примерно в 100 раз эффективнее Ca2+ в снижении негативного эффекта, индуцированного алюминием торможения удлинения корней у сои. Из этого следует, что отмеченный защитный эффект Mg2+ определяется изменениями не только поверхностного потенциала мембраны, а, возможно, обусловлен уменьшением связывания алюминия с G-белками.
Воздействие на животных. Если в 1970—1980 гг. можно было обнаружить 140 публикаций по теме «алюминиевая токсичность», то в 1990—2000 гг. — уже 1035. Интерес к данной теме особенно возрос после появления предположений о возможной связи алюминия и болезни Альцгеймера. Первые исследования были проведены на цыплятах. Симптомы токсичности включали: уменьшение потребления содержащего алюминий растительного корма, эффективности его усвоения и среднесуточного привеса тушки; нарушение обмена веществ и, в частности, уменьшение поглощения фосфатов, гиперкальцемию, пониженную минерализацию скелета и накопление алюминия в мышечной ткани. Поглощение большого количества растворимого алюминия (3—4 г/кг корма) может быть фатальным, особенно у молодых цыплят, если содержание кальция и фосфора в корме низкое. При этом интересно, что соединения, в частности окись алюминия, нерастворимые в разбавленных кислотах или воде, не обнаруживают симптомов токсичности вплоть до концентрации 16 г/кг корма. Вместе с тем растворимые соединения алюминия, например хлориды, сульфаты, ацетаты и нитраты, вызывают сильные симптомы токсичности уже при дозе 5 г/кг корма (в пересчете на Al).
Интересные данные, частично подтверждающие вышесказанное, получены при скармливании коровам Голынтинской породы в течение 7 нед. корма, содержащего 2 г/кг корма (в пересчете на Al) в виде растворимого хлорида алюминия. Потребление свежескошенной травы у опытных коров снизилось на 17%, суточный привес на 47%, а масса корма на 1 кг привеса, напротив, увеличилась на 50%. Одновременно содержание фосфора в фекалиях возросло на 79% при падении его уровня в плазме крови до соответствующего коровам, получающим корм с дефицитом фосфора. На пастбищах, в частности с посевом овсяницы, высокая концентрация Al (до 2 г /кг травы) вызывает у коров гиперкальциемию, которая в свою очередь приводит к потере магния с мочой. Алюминий накапливается в костном скелете, вызывая деминерализацию.
Каковы же возможные биохимические механизмы токсичности алюминия для животных? Прежде всего алюминий может влиять на белки, образуя межмолекулярные мостики с боковыми цепями аминокислот, изменяя скорость оборота белков, стабилизируя их или ускоряя деградацию благодаря конформационным изменениям.
Многие процессы передачи сигналов включают перенос фосфатных групп, и это звено может быть еще одним местом проявления токсичности алюминия. Как известно, сигнальная система с участием PIP2 ингибируется алюминием, видимо, путем его связывания с фосфатной группой PIP2 в мембранных фосфолипидах. Это в свою очередь подавляет гидролиз PIP2 фосфолипазой С. Изменение путей передачи сигналов может помочь объяснить изменение экспрессии генов в тканях в результате действия алюминия. Последний оказывает также заметное влияние на G-белки и протеинкиназы.
Одно из важных проявлений токсичности алюминия — это вмешательство в метаболизм железа за счет блокирования его включения в гем, приводящее к снижению концентрации гемоглобина и анемии. Алюминий нарушает также механизм контроля внутриклеточного гомеостаза железа, в результате чего может быть повышен уровень реактивного (свободного) железа и соответственно индуцирован продуцированный железом окислительный стресс.
Повышение концентрации свободного железа обычно стимулирует синтез железосодержащего запасного белка ферритина и ингибирует синтез рецепторов трансферрина, контролирующего поглощение железа. Алюминий противодействует эффектам внутриклеточного железа — регуляции трансляции мРНК рецепторов ферритина и трансферрина. В результате концентрация свободного железа в клетке относительно его запасных форм становится заметно выше, вызывая риск индуцированного им окислительного стресса. Алюминий может также ингибировать активность АТФ-зависимой протонной помпы на эндосомах, что приводит к удерживанию связанного в трансферрине железа внутри этих везикул. Это в свою очередь ограничивает активность железа в процессах синтеза ферритина.
Антиоксиданты снижают индуцированный алюминием окислительный стресс, что еще раз свидетельствует о важной роли алюминия в этом процессе.