



Токсическое действие тяжелых металлов на растения
Одной из важнейших черт металлиндуцированного изменения метаболизма клеток является, как правило, снижение активности ряда ферментов.
Активность ферментов. Разные металлы вызывают 50%-ную инактивацию большинства ферментов при разных молярных концентрациях: Ag+, Hg+, Cu2+ (10в-7—10в-5) > Cd2+ (10в-6 — 3*10в-5) > Zn2+ (10в-5 — 10в-4) >> Pb2+ (10-5 - 2*10в-4) > Ni2+ (10в-5 — 6*10в-4) > Co2+ 2(10-4 — 3*10в-4). Инактивация обусловлена взаимодействием металлов с SH-группами фермента, в результате чего изменяется его конформация. Кроме этого, Cd2+ и Pb2+ могут вытеснять Zn2+, связанный с SH-группами.
Загрязнение почвы никелем оказывало значительное влияние на ферментативную активность в растениях. Наблюдалось снижение активности каталазы, инвертазы (в 3 раза), щелочной фосфатазы (в 2,3 раза), и увеличение в 2,2 раза активности кислой фосфатазы.
Один и тот же фермент у разных видов (популяций) может быть в различной степени устойчив к воздействию тяжелых металлов. Карбоангидраза устойчивой к свинцу популяции перловника активировалась Pb2+, в то время как активность у неустойчивой популяции не повышалась из-за модификации молекулы фермента. Ион Cd2+ ингибировал in vivo Н+/К+-антипорт и АТФазную активность у растений в следующем порядке: капуста > тыква >> горох > рис > фасоль, что может быть отражением толерантности интактных растений.
Наряду с этим TM стимулируют образование в клетках активных форм кислорода. В ответ на окислительный стресс возрастает активность антиоксидантных ферментов (каталазы, пероксидазы, супероксиддисмутазы), что приводит к нейтрализации свободных радикалов и пероксидов. Вместе с тем активность антиоксидантных ферментов при кратковременном воздействии металлов может даже уменьшаться (рис. 11.1).

Устойчивость отдельных ферментов и активация ферментных систем, ответственных за стресс-метаболизм, может быть одной из возможных причин нечувствительности растений к избытку тяжелых металлов.
Рассмотрим ингибирующее действие TM на отдельные физиологические процессы.
Водный обмен. Изменение водного режима отрицательно сказывается на большинстве физиологических процессов в растениях — прорастании семян, фотосинтезе и дыхании, росте и развитии. Поэтому нарушение параметров водного обмена (в частности, изменение относительного содержания воды в тканях, водного потенциала и интенсивности транспирации) в присутствии высоких концентраций TM является одной из важных причин их токсичности.
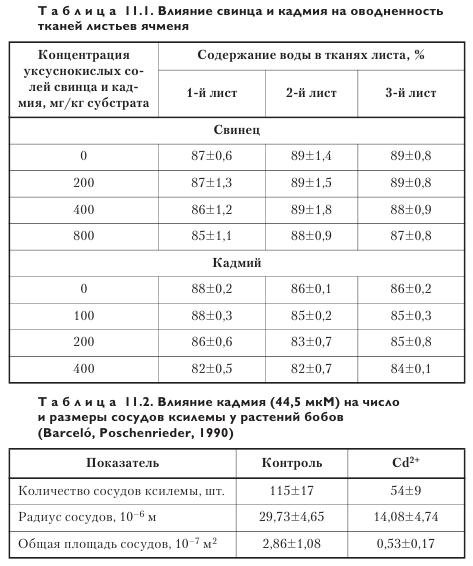
При низких концентрациях свинца и кадмия оводненность тканей листьев ячменя разного яруса практически не изменяется и остается на уровне, близком к контрольному варианту (табл. 11.1).
При повышении концентрации тяжелых металлов в корнеобитаемой среде содержание воды в клетках растений заметно снижается, что связано в первую очередь с уменьшением числа и диаметра сосудов ксилемы (табл. 11.2) и ситовидных трубок флоэмы. При очень высоких концентрациях металлов по указанной причине может сильно ограничиваться поступление воды, приводящее растение к гибели.
Уменьшение тургора растительных тканей в условиях повышенного содержания тяжелых металлов может быть также связано со снижением эластичности клеточных стенок сосудов, обусловленным частичным замещением кальция ионами металлов и изменением проницаемости мембран. Кроме того, уменьшение размеров корневой системы и числа корневых волосков снижает площадь всасывающей поверхности корня и, как следствие, содержание воды в клетках растений. Вместе с тем повышение количества связанной воды в этих условиях приводит к увеличению водоудерживающей способности тканей и снижению интенсивности транспирации, предотвращающих потерю воды.
В присутствии повышенных концентраций TM уменьшается водный потенциал растений, причем величина снижения в равной степени зависит от концентрации TM в корнеобитаемой среде и продолжительности обработки. Так, водный потенциал листьев фасоли в присутствии кадмия в концентрации 3 мкМ не изменялся в течение 120 ч обработки, но снижался от -0,23 МПа у контрольных до -0,37 МПа у опытных растений при более длительной экспозиции.
При обработке растений гороха, кукурузы и подсолнечника кадмием, а пшеницы никелем наблюдалось значительное снижение интенсивности транспирации, что может являться результатом закрытия устьиц. Под воздействием свинца в концентрациях 0,1 мМ и 1 мМ заметно снижалась устьичная проводимость у растений огурца. В присутствии Cd2+ и Ni2+ в концентрации 10 мМ у листьев пшеницы после 4-часовой обработки она составляла лишь 60—90% от контрольного варианта. Возможно это объясняется индуцированием ионами металлов изменений в регуляции К+-каналов в замыкающих клетках устьиц из-за резкого повышения уровня ABK и утечки ионов калия из клеток.
Помимо непосредственного воздействия TM на устьица, снижение потерь воды растением может быть связано с уменьшением размеров листьев и корневой системы, а также с нарушением поступления в замыкающие клетки ионов K+ и Ca2+.

В обобщенном виде влияние TM на некоторые показатели, связанные с водным обменом растений, отражено в таблице 11.3.
Таким образом, нарушение водного обмена у растений при действии TM обусловлено целым рядом причин:
• во-первых, уменьшением площади листьев;
• во-вторых, уменьшением размеров устьичных клеток и увеличением плотности их расположения за счет большего торможения роста листа, чем дифференцировки устьичных клеток;
• в-третьих, уменьшением содержания тургорогенов и пластичности клеточных стенок;
• в-четвертых, увеличением содержания АБК, индуцирующей закрывание устьиц.
Ион Cd2+, влияя на метаболизм АБК, стимулирует экспрессию в эпидерме ltp-тена, ответственного за синтез неспецифичных липидпереносящих белков. Это, в свою очередь, является причиной увеличения количества мономеров, поступающих к месту синтеза кутина и увеличения толщины кутикулы, затрудняющей транспирацию. Кроме того, нарушение дыхания и окислительного фосфорилирования также могут нарушать водный обмен растений.
В ответ на индуцированный металлом водный стресс наблюдается суперпродукция пролина, обладающего осморегуляторным, антиоксидантным и стресс-протекторным действием.
Фотосинтез. Фотосинтетический аппарат растений весьма чувствителен к повышенному содержанию TM в окружающей среде, что проявляется в изменении многих структурно-функциональных параметров фотосинтеза.
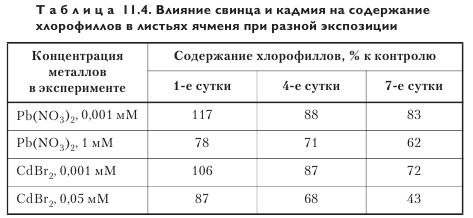
Пигменты фотосинтеза. Показано заметное уменьшение содержания хлорофиллов в листьях ячменя при действии высоких концентраций свинца и кадмия (табл. 11.4; рис. 11.2), причем с увеличением длительности обработки отрицательное влияние тяжелых металлов на содержание пигментов усиливалось. Степень отрицательного воздействия металлов на содержание пигментов зависит не только от токсичности металла, его концентрации и длительности обработки, но и от яруса листа. Так, в присутствии свинца и кадмия снижение содержания пигментов в 3-м листе ячменя было выражено сильнее, чем в 1-м и 2-м.

Вместе с тем соотношение хлорофиллов а и b сохраняется практически неизменным в присутствии даже высоких концентраций металлов. Известно, что хлорофилл а входит в состав реакционных центров и периферических антенных комплексов ФС1иФСП, в то время как хлорофилл b преимущественно компонент светособирающего комплекса ФС II. Постоянство отношения хлорофиллов а и b указывает на неизменность стехиометрического соотношения между комплексами реакционных центров фотосистем I и II и светособирающего комплекса ФС II.
Главной причиной снижения содержания зеленых пигментов в присутствии TM является подавление биосинтеза хлорофилла, связанное в первую очередь с непосредственным действием металлов на активность ферментов биосинтеза. Основными точками ингибирования при этом выступают образование фотоактивного хлорофиллидредуктазного комплекса и синтез b-аминолевулиновой кислоты (рис. 11.3). Опосредованное действие металлов на биосинтез хлорофилла связано, как полагают, с дефицитом железа. Кроме того, нарушение биосинтеза хлорофилла в присутствии TM может быть вызвано вытеснением этими ионами Mg2+ из молекулы хлорофилла. Некоторые металлы, например медь, в больших концентрациях замедляют связывание молекул хлорофиллов с белками в светособирающих комплексах фотосистем.
Каротиноиды по сравнению с хлорофиллами менее подвержены негативному действию TM, что, видимо, связано с выполняемой ими защитной ролью.

Ультраструктура хлоропластов. В мембранах хлоропластов обнаружены Zn/Cd-белки-переносчики, что говорит о возможности попадания TM в эти органеллы и, следовательно, их непосредственном воздействии на ультраструктурную организацию хлоропластов. В частности, в присутствии высоких концентраций кадмия изменяется структура мембран тилакоидов, что может быть результатом гидролиза липидов и освобождения жирных кислот. Одновременно в результате усиления деградации органелл увеличивается количество пластоглобул, уменьшается количество гран и нарушается их структура (граны содержат меньшее количество тилакоидов), также наблюдается деградация ламелл стромы (табл. 11.5).

Отмеченные нарушения вызывают инактивацию кислородвыделяющего центра ФС II и замедление электронного транспорта.
Световая фаза фотосинтеза. Наиболее чувствительна к ионам металлов ФС II, активность которой оценивается на основании анализа кинетики замедленной флуоресценции хлорофилла. Для возникновения замедленной флуоресценции необходим интактный фотосинтетический аппарат (ФСА). Замедленную флуоресценцию используют также в качестве критерия при оценке его целостности. Изменения в присутствии TM таких параметров флуоресценции хлорофилла, как максимальный (Fm) и минимальный (F0) выход флуоресценции, переменная флуоресценция (Fv), а также квантовая эффективность ФС II (Fv/Fm), указывают на определенные нарушения в ФСА растений.
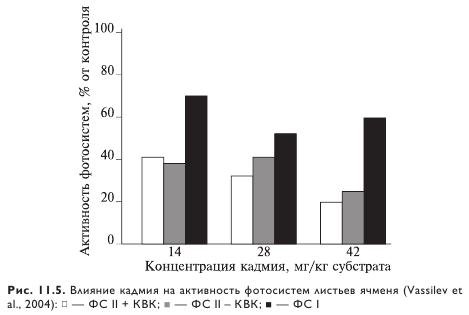
Достоверное уменьшение эффективности ФС II у однолетних злаков, в частности Setaria viridis, наблюдается у растений лишь при использовании самой высокой из изученных концентрации кадмия (160 мг/кг субстрата). У ячменя даже эта концентрация не вызывает изменений (рис. 11.4, 11.5). Установлено также некоторое снижение фотохимического тушения (qP) и одновременно повышение уровня нефотохимического тушения (qN). Аналогичную реакцию ФСА в присутствии кадмия наблюдали у растений бобов, пшеницы, подсолнечника и ячменя. Уменьшение эффективности ФС II отмечено также у растений бобов при действии повышенных концентраций меди. Сильное увеличение qN может быть результатом угнетения активности цикла Кальвина, а также нарушения утилизации фотосинтетической энергии. Уменьшение же qP связывают с нарушениями в цепи транспорта электронов.

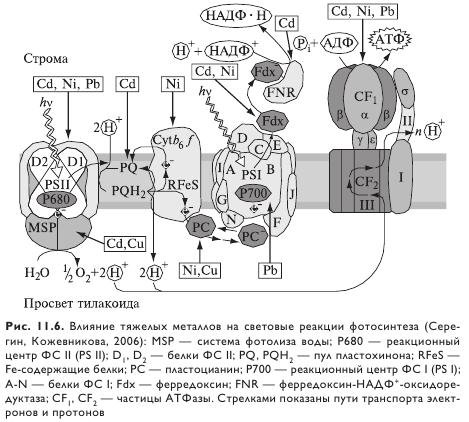
Предполагается, что наиболее важными причинами снижения активности ФСП в присутствии повышенных концентраций TM являются изменение структуры белков реакционного центра и взаимодействие ионов некоторых металлов (Cd2+, Zn2+) с ионами, входящими в состав кислородвыделяющего центра (Mn2+, Ca2+, Cl-). Мишенью действия TM, в частности кадмия, служит первичный донор электронов реакционного центра ФС II — П-680. Имеются сведения и о повреждении TM светособирающих антенных комплексов реакционных центров фотосистем I и II. Хотя ФС I в целом считается относительно устойчивой к действию TM, некоторые нарушения обнаружены и здесь, в частности это подавление работы ФС I медью, обусловленное взаимодействием металла с ферредоксином, а также изменениями в количестве пластоцианина, содержащего атомы меди.
Тяжелые металлы непосредственно влияют на перенос электронов в фотохимических реакциях. Так, кадмий в концентрациях 14, 28 и 42 мг/кг субстрата снижал скорость электронного транспорта в мембране тилакоидов растений ячменя. У растений люцерны, клевера и бобов кадмий блокировал транспорт электронов и протонов на уровне пластохинона при переходе электронов от ФС II к ФС I.
Возможная причина ингибирования TM фотосинтетического электронного транспорта — изменения в ультраструктуре хлоропластов, в частности повреждение тилакоидов. Кроме того, нарушения в переносе электронов могут быть связаны с вызываемым тяжелыми металлами дефицитом железа.
Тяжелые металлы оказывают определенное негативное действие и на реакции фотофосфорилирования. Так, у обработанных кадмием растений ячменя замедление циклического и нециклического фотофосфорилирования наблюдается уже при концентрациях металла в растворе 1,7 мкМ. Снижение фотофосфорилирования обнаружено в присутствии свинца в концентрации 20 мкМ и меди в концентрации 1,7 мкМ.
Воздействие TM на световую фазу фотосинтеза показано на схеме (рис. 11.6).
Темновая фаза фотосинтеза. Основными «мишенями» токсического действия TM служат ферменты цикла Кальвина. Так, инактивация кадмием РБФК может быть вызвана нарушением ее структуры в результате взаимодействия TM с SH-группами (рис. 11.7). Кроме того, кадмий и цинк уменьшают активность РБФК, замещая Mg2+ в молекуле фермента. Имеет также место ингибирование синтеза некоторых ферментов цикла Кальвина de novo, например глицеральдегид-3-фосфатдегидрогеназы.

Замедление скорости реакций темновой фазы фотосинтеза в присутствии TM может быть обусловлено также снижением поступления в клетки CO2 из-за уменьшения числа устьиц или их закрывания. Так, даже кратковременное (3 ч) воздействие нитратом свинца в концентрациях 0,1 и 1 ммоль на корневую систему проростков огурца вызывает снижение нетто-фотосинтеза и одновременное уменьшение устьичной проводимости и транспирации (табл. 11.6). Вызывая быстрое закрывание устьиц, свинец ограничивает поступление CO2 в лист и, соответственно, уменьшает скорость фотосинтеза.

Вместе с тем при определенных концентрациях TM могут оказывать стимулирующее действие на фотосинтез. В частности, показано увеличение содержания хлорофиллов в листьях кукурузы и ячменя в присутствии цинка и свинца в концентрациях 1 ммоль. При использовании цинка в концентрациях 5— 20 ммолей наблюдалось увеличение активности РБФК у растений Lolium perenne L. Кратковременное (3 ч) действие нитрата свинца в концентрации 0,01 ммоль приводило к повышению интенсивности фотосинтеза у проростков огурца (см. табл. 11.6). Стимуляция фотосинтеза ацетатом свинца (200 мг/кг субстрата) в вегетационных опытах отмечена у растений овса и ячменя. Возможно, что усиление фотосинтетических процессов связано с увеличением активности ФС II, а также обусловлено общей активизацией метаболизма растений в ответ на слабое действие стрессора.
Таким образом, тяжелые металлы могут воздействовать на фотосинтез, уменьшая содержание фотосинтетических пигментов и активность ферментов цикла Кальвина, нарушая ультраструктуру хлоропластов, а также замедляя скорость электронного транспорта. Степень ингибирования фотосинтеза во многом зависит от концентрации металла в субстрате. Высокие концентрации металлов снижают интенсивность процесса ассимиляции CO2 как за счет их прямого действия на отдельные реакции фотосинтеза, так и в результате опосредованного влияния на другие физиологические процессы.
Дыхание. Устойчивость дыхания растений к повышенному содержанию TM в целом достаточно высока, и в довольно широком диапазоне концентраций они не вызывают у растений заметных изменений в интенсивности дыхания. Дальнейшее повышение концентрации TM в окружающей среде приводит к усилению дыхания, и лишь концентрации, близкие к пороговым, замедляют его. Например, в листьях гороха, ячменя и кукурузы интенсивность дыхания под влиянием свинца в концентрации 5 ммоль увеличивалась на 20—50% по отношению к контролю. При воздействии кадмия в концентрации 0,9 и 1,35 ммоль наблюдалось усиление дыхания в листьях бобов, а в присутствии ртути — в подземных и надземных органах редиса, салата и ячменя. В присутствии уксуснокислой соли свинца в концентрациях 200, 400 и 800 мг/кг субстрата и кадмия в концентрациях 100 и 200 мг/кг субстрата не было обнаружено изменений в интенсивности дыхания у растений ячменя. Усиление дыхания наблюдается у растений также при увеличении продолжительности обработки металлом.
Основными причинами усиления интенсивности дыхания являются активация некоторых ферментов, увеличение энергетических затрат организма на процессы поддержания и репарации повреждений в клетках, вызванных действием стресс-фактора, а также необходимость увеличения синтеза органических кислот (малата, цитрата, сукцината), являющихся хелаторами TM.
Гликолиз, цикл Кребса и пентозофосфатный путь окисления глюкозы. Замедление скорости дыхания в первую очередь связано с изменением активности дыхательных ферментов. Например, на проростках гороха и бобов показано ингибирование кадмием в концентрации 1 ммоль активности ферментов гликолиза (в частности гексокиназы) и пентозофосфатного пути (глюкозо-6-фосфатдегидрогеназы, 6-фосфоглюконатдегидрогеназы). Это объясняется изменениями в структуре ферментов, а также замедлением их синтеза de novo. Вместе с тем показана возможность активации пентозофосфатного цикла в условиях пониженного содержания тяжелых металлов, связанной, очевидно, с увеличением активности ключевого фермента — глюкозо-6-фосфатдегидрогеназы.
Снижение активности других ферментов, в частности малатдегидрогеназы и изоцитратдегидрогеназы, являющееся одной из основных причин уменьшения ИД в условиях повышенного содержания TM в субстрате, объясняется непосредственным взаимодействием ионов металлов с SH-группами молекул белков.
Уменьшение интенсивности дыхания может быть также обусловлено нарушением проницаемости мембран митохондрий. Первичным звеном в митохондриях, на которое воздействуют металлы, видимо, является сукцинатдегидрогеназный комплекс.
Замедление скорости дыхания растений в присутствии высоких концентраций TM может быть связано также с ингибированием транспорта электронов и протонов в митохондриях, в результате чего нарушается нормальное функционирование транспортной цепи электрон. Наблюдается также разобщение окислительного фосфорилирования за счет увеличения пассивной проницаемости для протонов внутренней мембраны митохондрий.
Таким образом, влияние TM на дыхание в большой степени зависит от силы воздействия. Так, умеренный стресс, как правило, не сказывается на интенсивности дыхания растений или может увеличивать его, тогда как высокие концентрации заметно снижают скорость этого процесса.
Минеральное питание. Между тяжелыми металлами и макроэлементами возможны как антагонистические, так и синергические взаимодействия (табл. 11.7).

Влияние тяжелых металлов на содержание макроэлементов. Азот. Высокие концентрации TM, в частности меди, снижали концентрацию азота в листьях растений пшеницы. В присутствии цинка его содержание уменьшалось в надземной биомассе растений овса, а в присутствии кадмия — в корнях и листьях бобов. Негативное влияние TM на азотный метаболизм связано с конкуренцией за мембранные переносчики между ионами металлов и NH4+, а также со снижением активности некоторых ферментов азотного метаболизма: нитритредуктазы, глутаматсинтазы и аминотрансферазы в листьях, нитратредуктазы в корнях.
Относительно низкие концентрации TM могут вызывать увеличение содержания азота в растениях, что связано с активацией ферментов. Так, у растений овса кадмий в концентрации 100 мкМ на 15% (по сравнению с контролем) увеличивал активность глутаминсинтетазы, участвующей в ассимиляции аммония, что может способствовать повышению устойчивости азотного метаболизма.
Фосфор. В зависимости от вида металла и его концентрации содержание фосфора может как уменьшаться, так и увеличиваться. Так, в присутствии относительно невысоких концентраций кадмия повышалось его содержание в корнях и листьях растений Sedum alfredii, а при действии цинка — в стеблях овса и в листьях и корнях ячменя. При этом повышение содержания фосфора в присутствии цинка можно объяснить синергическим взаимоотношением этих элементов, а причина его увеличения под влиянием других TM пока остается неизвестной. Более высокие концентрации кадмия приводили к снижению содержания фосфора в надземной массе арабидопсиса, а высокие концентрации меди — в листьях овса. Возможно это связано с нарушением поглощения и передвижения элемента в растениях.
Калий и кальций. Характер изменения содержания в растениях калия и кальция в присутствии TM во многом зависит от видовой принадлежности, а также органа растения. Строгой зависимости между содержанием TM в субстрате и концентрацией калия и кальция в органах растений не выявлено. К примеру, при повышении в субстрате концентрации кадмия до 1 ммоля содержание калия в листьях Sedum alfredii увеличивалось, тогда как в листьях пшеницы уменьшалось. У растений бобов под действием высоких концентраций меди содержание кальция в корнях повышалось, а в листьях понижалось, в то время как содержание калия в листьях, наоборот, увеличивалось. Известны антагонистические отношения между кальцием и свинцом, связанные с конкуренцией за участки поглощения.
Увеличение содержания K+ и Ca2+ при возрастании содержания TM в среде, очевидно, играет защитную роль, однако механизм этого явления пока неизвестен. Снижение же содержания калия и кальция в клетках в большей степени связано с нарушениями структуры клеточных мембран и утечкой этих ионов из клеток.
Магний. Сведений относительно влияния TM на содержание магния в растениях очень мало. Отмечено, в частности, что в присутствии кадмия уменьшалось содержание магния в листьях ячменя, а при действии свинца — в листьях огурца. Вместе с тем при определенных концентрациях TM содержание Mg2+ в органах может и повышаться. Например, в присутствии цинка в концентрациях 0,4 и 1,6 ммоль/л питательного раствора содержание магния в корнях ячменя снижалось на 70 и 87% по отношению к контролю соответственно, тогда как в листьях несколько повышалось — на 6 и 17% соответственно. Обнаружено также увеличение содержания магния в надземных органах растений овса под влиянием меди.
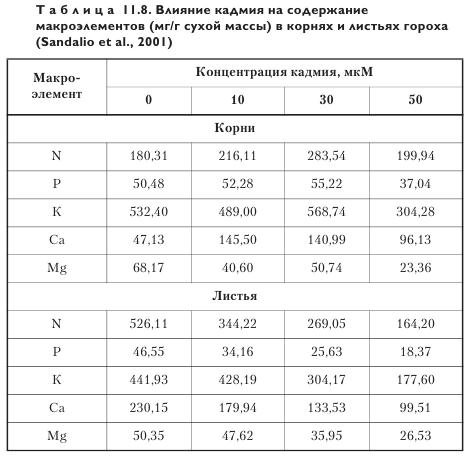
Возможные изменения количества макроэлементов в разных органах растений в присутствии TM, в частности кадмия, показаны на примере гороха (табл. 11.8). Содержание макроэлементов в листьях по мере увеличения концентрации кадмия в корнеобитаемой среде снижается, а в корнях отмечено как повышение, так и снижение.
Влияние тяжелых металлов на содержание некоторых микроэлементов. Железо. Известно, что железо и цинк являются антагонистами, и катионы цинка замедляют поглощение железа, вызывая хлороз листьев. Сходный эффект, вызываемый кадмием, выражается в уменьшении размеров корневой системы, повреждении клеточных мембран корня, задержке транспорта железа из корней в листья, а также снижении активности Fe-peдуктазы — фермента, необходимого для восстановления Fe3+ в Fe2+. Уменьшение содержания железа отмечено также в листьях растений огурца под действием свинца и меди (табл. 11.9).

Марганец, медь, цинк, бор. Помимо железа TM нарушают в растениях метаболизм других микроэлементов. Так, с увеличением концентрации кадмия уменьшается содержание марганца, меди и бора в листьях ячменя. У растений салата при действии кадмия в концентрации 1 мг/л содержание марганца в стебле увеличивалось, а в корнях уменьшалось. В присутствии кадмия в концентрации 10—30 мкмоль также снижалось поступление марганца в растения пекинской капусты и гороха, что в свою очередь приводило к снижению количества гран в хлоропластах и тилакоидов в гранах.
При действии цинка даже в относительно невысоких концентрациях (100 мкМ) уменьшается концентрация марганца и меди в корнях и надземной массе растений арабидопсиса.
Свинец подавляет поглощение и транспорт марганца в побеги пшеницы, нарушая физические процессы связывания ионов.
Макро- и микроэлементы в свою очередь также могут снижать концентрацию TM в растении. Так, увеличение содержания калия и фосфора является причиной уменьшения поглощения кадмия. Отмечено падение уровня свинца и кадмия в растениях риса при повышении в почве содержания кальция и магния. Наконец, и сами TM ограничивают поступление друг друга в растения. Так, цинк снижает поступление свинца, а медь и железо — кадмия.
Поглощение ионов. В большинстве случаев кадмий и свинец тормозят поглощение как катионов (К+, Ca2+, Mg2+, Mn2+, Zn2+, Cu2+, Fe3+), так и анионов (NO3).
Для некоторых элементов картина может быть более сложной. Накопление меди снижалось при воздействии кадмия в корнях райграса, кукурузы, капусты, клевера, но увеличивалось в корнях риса и оставалось на прежнем уровне у огурца и тыквы. Тяжелые металлы влияют на поглощение других ионов посредством разных механизмов, относительная роль которых неодинакова, поэтому эффекты у отдельных видов могут отличаться.
Можно выделить по крайней мере два механизма снижения поглощения макро- и микроэлементов под воздействием Cd2+ и Pb2+. Первый определяется физико-химическими причинами и обусловлен близостью ионных радиусов металлов. Таким образом, например, Cd2+ (1,03 А) снижает поглощение Zn2+ (0,83 А) и Ca2+ (1,06 А). Второй механизм связан с металл-индуцированным нарушением метаболизма клеток, в результате чего происходят изменение активности мембранных ферментов и структурная перестройка мембран. Одним из самых ярких примеров подобного действия Cd2+ и Pb2+ является вызываемый ими отток К+ из корней. Вероятно, К+-АТФаза и SH-группы белков клеточной мембраны очень чувствительны к Cd2+ и Pb2+, что и является причиной оттока К+ из корней, одного из типичных процессов, наблюдаемых при самых разных нарушениях метаболизма клеток. С металл-индуцированным снижением активности другого фермента — Fе(Ш)-редуктазы связано уменьшение поглощения Fe3+ корнями огурца.
Под действием Cd2+ происходило резкое изменение липидного состава мембран, причем во всех классах липидов содержание пальмитиновой кислоты увеличивалось, а линолевой и линоленовой кислот, напротив, уменьшалось. Следствием этого могло быть изменение проницаемости мембран, что, наряду с ингибированием мембранных ферментов, возможно, приводило к изменению ионного баланса в цитоплазме.
Поглощение NO3 уменьшалось при действии Pb2+ и Cd2+, что приводит к уменьшению активности нитратредуктазы и нарушению азотного обмена растений. Снижение поглощения NO3 может быть обусловлено также водным стрессом, вызываемым Cd2+ и Pb2+.
Неоднозначно изменялся ионный баланс в различных тканях и органах растений. Так, Pb2+ вызывал уменьшение содержания К+ во всех органах проростков огурца, Ca2+ в гипокотиле и семядолях, Fe3+ только в корнях. Ион Cd2+ понижал содержание К+ в корнях и семядолях, Ca2+ только в семядолях, а Fe3+ только в корнях. При комбинированном действии Cd2+ и Pb2+ чаще наблюдался синергизм, чем антагонизм.
TM влияют не только на поглощение ионов, но и на их транспорт. Это влияние видоспецифично и, возможно, является одной из причин различной чувствительности к ним растений. Например, кадмий уменьшал как поглощение, так и транспорт Zn2+, Fe3+, Mn2+, Ca2+ и Mg2+, причем в большей степени у клевера, чем у капусты.
Рост и морфогенез. Токсическое действие металлов четко видно по ингибированию роста, что широко применяется для тестирования их присутствия в окружающей среде. Торможение роста происходит, с одной стороны, из-за нарушения метаболизма и, с другой — в результате прямого действия металлов на рост, например в результате взаимодействия с полисахаридами клеточных стенок и снижения их пластичности.
Нaибoлee устойчиво к TM прорастание семян, что обусловлено низкой проницаемостью для них семенной кожуры большинства видов. У видов, семенная кожура которых хорошо проницаема для Pb2+ (например, Phaseolus vulgaris, Pisum sativum), семена теряли способность прорастать в присутствии солей свинца. Ион Cd2+ в малых концентрациях (10в-5 М) может даже стимулировать прорастание семян.
Рост корня более чувствителен к TM, чем рост побега, что обусловлено их преимущественным накоплением в корнях. Токсичность металлов уменьшалась в следующем порядке (при расчете концентрации в г/л): Cu = Tl > Ag > Cd > Hg > Co > Zn > Pb.
Линейное уменьшение роста корня проростков ячменя и кукурузы наблюдали при концентрации свинца 3750—12 371 мг/кг сухой массы для ячменя и 1722—4828 мг/кг сухой массы для кукурузы. Снижение роста корней ячменя на 10% начиналось при содержании свинца примерно 525 мг/кг сухой массы, а кукурузы — 183 мг/кг сухой массы.
Диапазон концентраций как Cd2+, так и Pb2+, при которых они ингибируют рост, достаточно узкий. Так, например, для корней проростков кукурузы летальные концентрации были примерно на порядок больше тех, при которых начиналось достоверное ингибирование их роста.
TM вызывают изменение морфологии корней. Если концентрации металлов не слишком высоки, число боковых корней уменьшается в меньшей степени, чем длина главного корня, и корневая система имеет более компактный вид. Процесс заложения боковых корней очень устойчив к воздействию TM (за исключением Ni), что может быть обусловлено как барьерной ролью эндодермы, так и структурными особенностями клеток центрального цилиндра.
Снижение скоростей как деления, так и растяжения клеток под действием TM происходит за счет разных механизмов: прямого связывания с ДНК, металл-индуцированных аберраций, удлинения митотического цикла, нарушения формирования микротрубочек, снижения пластичности клеточных стенок, уменьшения пула глутатиона.
Специфика реакций разных видов растений на действие TM определяется в значительной мере способностью к синтезу металлсвязывающих соединений и выведению поглощаемых металлов из активного метаболизма. Немаловажно, в какой культуре проводился опыт — водной, песчаной или почвенной (табл. 11.10).

Из таблицы видно, во-первых, отсутствие связи между концентрацией кадмия в питательном растворе и его содержанием в листьях и, во-вторых, ранжирование культур по чувствительности к Cd2+ в песчаной и водной культуре. Концентрации вплоть до 0,10 мг/л понижают на 25% накопление биомассы свеклы, фасоли, турнепса, тогда как для такой же задержки роста капусты требуется 4 мг/л кадмия.
Бингам и сотр выращивали ряд сельскохозяйственных культур в сосудах на супеси при pH = 7,5, используя городские отходы с различным количеством CdSO4, причем отходы вносились в количестве 10 мг/кг почвы, а растения выращивались до хозяйственной спелости. Оказалось, что добавления 10 мг кадмия на 1 кг почвы уже достаточно, чтобы понизить урожай шпината, соевых бобов и кресс-салата на 25% или более. Напротив, концентрации кадмия, превышающие 160 мг/кг почвы, требовались для такого же понижения урожайности томатов, огурцов, капусты, свеклы и риса.
Из приведенных данных понятно, что окончательные выводы о реакции растений на TM можно сделать по итогам вегетационных опытов в почвенной культуре или, что еще лучше, микрополевых опытов.