



Затопление и засоление: причины, взаимообусловленность и последствия для растений
Большинство засоленных почв в мире подвержены затоплению (насыщению почвы водой) из-за наличия неглубокого водного зеркала или уменьшенной инфильтрации поверхностных вод из-за плохих физико-механических свойств почв. Гипоксия (низкая концентрация O2) в почве возникает в результате низкой растворимости O2 в воде (0,28 моль/м3 при 20 0C), низкой диффузности O2 в заполненных водой порах (примерно в 100 000 раз медленнее, чем через воздухоносные почвенные поры), а также быстрого использования растворенного O2 бактериями и корнями.
Причины повышенной концентрации ионов. Концентрация ионов в листьях представляет собой результат поступления ионных потоков в листья и эффекта разбавления из-за накопления биомассы при росте (табл. 10.2). Возникает естественный вопрос, что является причиной повышения концентрации Na+ и Cl- — уменьшение скорости роста побегов или увеличение скорости ионного транспорта?
У пшеницы OCP побегов за первые четыре дня после начала гипоксии в расчете на сухую массу составила 0,24 сут-1 (несколько выше, чем для аэрируемого контроля), a OCP корней — 0,05 сут-1, что на 70% ниже, чем в контроле. В интервале 4—8-й день после начала гипоксии скорость роста снизилась до 0,13 сут-1, а рост корней прекратился совсем. Можно заключить, что повышенные концентрации Na+ и Cl- в побегах в результате затопления могли бы в принципе быть обусловлены пониженной скоростью роста. Для проверки этого положения были проведены специальные расчеты, причем в качестве исходных взяты данные из таблицы 10.2.
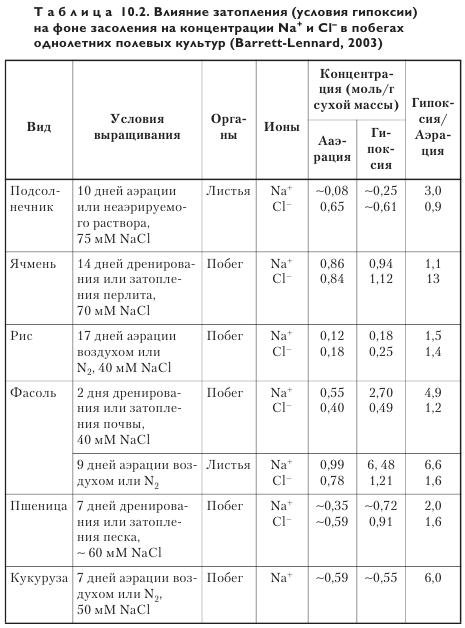
При затоплении (условия гипоксии) кукурузы и пшеницы на 7 дней вклад снижения скорости роста в повышение концентрации ионов колебался от 1 до 20%, а при более длительном затоплении ячменя и риса (14—17 дней) — 40—100%. В целом эти данные согласуются с точкой зрения, что кроме ингибирования скорости роста есть другая причина повышения концентрации ионов, что и подтвердилось в опытах на фасоли. При затоплении 17-дневных растений, выращиваемых при 40 мМ NaCl, на 1—3 дня уже после первых суток концентрация Na+ в побеге повысилась на 74%. К сожалению, масса побега в данной работе не определялась. Однако если бы даже затопление вызвало полную остановку роста и скорость транспорта Na+ оставалась бы постоянной, концентрация Na+ через сутки затопления увеличилась бы только на 13%. Значит, имеется какой-то другой фактор, играющий важную роль в этом процессе. Измерения скорости транспорта ионов за короткий интервал времени (1 сут) с помощью радиоактивных изотопов, а при более продолжительном периоде — посредством анализа содержания ионов в побеге показали, что затопление кукурузы и ячменя, выращиваемых при 50—60 мМ NaCl, на период 1—7 сут увеличило скорость транспорта Na+ и/или Cl- на 100—780%. При сходном засолении, но более продолжительном затоплении (14—17 сут) у ячменя и риса наблюдалось умеренное увеличение скорости нетто-транспорта Na+ и Cl- при одновременном заметном развитии аэренхимы в корнях, способствующей диффузии O2. Интересно, что 7-суточная гипоксия растений галофита Atriplex amnicola, выращиваемого при 400 мМ NaCl, не оказала влияния на нетто-накопление Na+ и Cl-.
Таким образом, повышенные концентрации Na+ и/или Cl- в побегах при засолении/затоплении лишь частично обусловлены снижением скорости их роста. Повышение скорости транспорта Na+ и Cl- также может быть причастно к этому событию.
Причины повышения скорости транспорта. Затопление (условия гипоксии) влияет на избирательность мембран, транспорт питательных веществ и удержание ионов и метаболитов с малой молекулярной массой корнями. Можно предположить наличие двух типов нарушения мембранных барьеров или процессов:
• значительная потеря мембранной целостности, в результате чего транспирация приводит к поглощению и транспорту ионов по схеме массового тока;
• более специфическое воздействие на поглощение и выход ионов, обусловленное дефицитом АТФ.
Если анализировать первую возможность, то можно было бы ожидать транспорт Na+, K+ и Cl- в побег в соотношениях, аналогичных тем, что имеют место в питательном растворе. Более того, корни были бы хорошо проницаемы и имели бы низкую концентрацию ионов после промывания свободного пространства корня. Однако результаты опытов показывают, что, несмотря на сочетание засоления и гипоксии, корни сохраняют удивительную способность регулировать (по меньше мере частично) содержание ионов и их транспорт в побег. Повышенная концентрация Na+ и/или Cl- в побегах затопленных растений при засолении, очевидно, обусловлена действием более тонких механизмов, чем просто резкая потеря целостности мембран корня.
Каким же образом затопление действует на функции корней, повышая скорости нетто-транспорта ионов? Помочь в получении ответа на данный вопрос могло бы определение влияния гипоксии и затопления на концентрации Na+ и Cl- в ксилеме и клетках стели. Показательно сравнение концентраций Cl- в ксилеме у ячменя — гликофита, выращиваемого при 50 мМ NaCl, и галофита Aricennia marina, выращиваемого при 400 мМ NaCl. Они составляли соответственно 2—5 и 5—37 мМ Cl-. Видимо, повышенные скорости транспорта Na+ и Cl- в побеге обусловлены повышенной концентрацией Na+ и Cl- в ксилеме. Однако это требует дополнительных подтверждений.
Наконец, любое объяснение на клеточном уровне механизма, с помощью которого гипоксия увеличивает поступление Na+ и Cl- в корни, должно согласоваться с современным представлением о регуляторной роли ионных каналов, трансмембранном градиенте протонов и Н+/АТФазах. в этой связи важны два аспекта проблемы. Во-первых, концентрации Na+ и Cl- в зоне загрузки ксилемы в корнях должны, видимо, в значительной мере регулироваться трансмембранным градиентом протонов в плазмалемме клеток коры. Во-вторых, эти протонные градиенты поддерживаются активностью Н+/АТФазы, видимо, поэтому дефицит АТФ, индуцируемый затоплением/гипоксией, влияет на них отрицательно.
Возможные физиологические адаптации при сочетании затопления и засоления. Растения используют несколько стратегий для улучшения регуляции транспорта солей при сочетании затопления и засоления:
• во-первых, формируют аэренхиму в тканях, чтобы избежать гипоксии, и регулируют поглощение и транспорт ионов посредством формирования эндодермы;
• во-вторых, снижают устьичную проводимость;
• в-третьих, защищают метаболизм путем удаления избытка солей.
Образование эндодермы и аэренхимы. Предварительное выдерживание растений в условиях гипоксии улучшает способность адаптироваться к засолению в сочетании с затоплением (табл. 10.3).
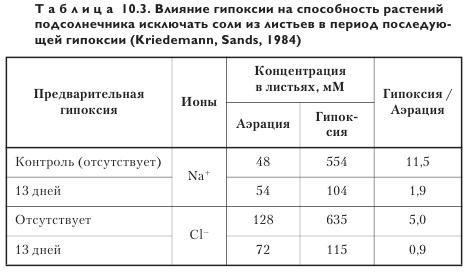
Предварительное воздействие на выращиваемые без засоления растения 13-дневной гипоксией вызывало незначительное (1,5—2 раза) повышение концентраций Na+ и Cl- в листьях в варианте гипоксия + засоление в сравнении с вариантом аэрация + засоление. В контроле (без предварительной гипоксии) это повышение для Na+ и Cl- составило соответственно 11 и 5 раз.
Каковы же механизмы, индуцируемые в растениях подсолнечника предобработкой гипоксией и запускающие систему ионной регуляции при сочетании засоления с затоплением? Самые значимые из них — развитие большого числа придаточных корней, формирование аэренхимы в коровых клетках гипокотиля и корней, формирование мощного эндодермального слоя.
Однако сразу же возникает вопрос — если аэренхима является таким важным атрибутом растений засоленных почв, почему она так не конститутивна, т. е. не присутствует в них постоянно? Большинство видов в ответ на гипоксию увеличивает формирование аэренхимы. В эволюционном смысле наличие конститутивной аэренхимы, видимо, делает растения менее конкурентоспособными, чем те из них, которые образуют ее в ответ на стресс.
Снижение устъичной проводимости. Затопление может вызвать закрывание устьиц в варианте как без засоления, так и с засолением. Закрывание устьиц будет выгодно растению в условиях засоления, если неожиданное засоление вызовет резкое повышение концентрации Na+ и Cl- в ксилеме. Семидневная гипоксия на фоне сильного засоления (400 мМ NaCl) вызывала удвоение концентрации Cl- в ксилеме у галофита Atriplex amnicola, а 14-дневная — утроение концентрации с последующим снижением устьичной проводимости на 70% по мере дальнейшего увеличения периода действия гипоксии. Вполне возможно, что растения противодействуют закрытием устьиц повышению концентрации Cl" в ксилеме и уменьшению скорости тока веществ в побег по ксилеме. Однако эта гипотеза пока не проверена на сельскохозяйственных культурах-гликофитах.
Защита метаболизма путем удаления солей. Гликофиты могут защищать листья путем удаления Na+ и Cl- из ксилемного тока и локализации их в клетках стебля, черешков и листовых влагалищ. Однако эти зоны насыщаются при концентрации Na+ около 1—3 мМ/г сухой массы. Их способность регулировать ионные потоки в ксилеме затопленных растений при засолении зависит от скорости роста, доли указанных органов в сухой массе побега (стебель фасоли ~ 35%, листовые влагалища ячменя ~ 25%) и скорости движения ионов по ксилеме после этих зон. Если принять во внимание увеличение концентрации Na+ на 560% в листьях фасоли, подвергнутых 9-суточной гипоксии при 40 мМ NaCl, можно заключить, что эти зоны играют ограниченную роль в контроле движения Na+ к листьям в этих условиях.
Таким образом, лучшим путем адаптации растений к условиям одновременного затопления и засоления может быть сочетание стратегий, включающих поддержание избирательной проницаемости мембран и снижение устьичной проводимости непосредственно после затопления при одновременном формировании новых придаточных корней, содержащих по возможности аэренхиму. Однако для окончательного выяснения механизмов требуются дальнейшие опыты, в которых определялись бы параметры роста, ионный статус (концентрации Na+ и Cl- в корнях, побеге, ксилемном соке и скорость транспорта ионов из корня в побег), скорость формирования аэренхимы и параметры водного режима. Интересными при этом могут быть опыты с близкородственными генотипами, но отличающимися устойчивостью к действию изучаемых факторов. Так, у сорта кукурузы LG11 гипоксия в течение 24 ч увеличила скорость транспорта 22Na+ в побег при 2,4 мМ NaCl — на 225%, при 10,9 мМ NaCl — на 38% и при 40 мМ NaCl не оказывала влияния. Вместе с тем у сорта Пионер 3906 установлено увеличение указанной скорости на 350—630% в диапазоне 0,2—50 мМ NaCl.
Влияние последействия взаимодействия затопления и засоления на ткани листьев и выживаемость растений. Затопление (условия гипоксии) на фоне засоления ускоряет старение листьев, причем старые листья пшеницы и кабачков, характеризующиеся накоплением большего количества Na+ и Cl-, отмирали первыми. В этом опыте при засолении выше 30 мМ NaCl скорость старения листьев превышала скорость появления новых, а рост побегов полностью ингибировался. Установлена следующая временная последовательность процессов по чувствительности к совместному действию засоления и затопления:
• после 1-й нед. действия факторов прекратился рост корней;
• после 2-й нед. началось отмирание наиболее старых листьев на главном побеге;
• после 3-й нед. скорости появления новых листьев и отмирания старых (-0,12—0,15 листьев/сут) уравнялись;
• через 5 нед. прекратился рост побегов, при этом формирование зерна было проблематичным.
В отсутствие засоления, но при действии гипоксии растения выглядят здоровыми, однако накопление биомассы снижается на 60% по сравнению с условиями аэрации. При 20 мМ NaCl и гипоксии растения выглядят хлоротичными, их масса на 60% ниже, чем в аэрируемом варианте. При 120 мМ NaCl побеги при гипоксии погибали, а при аэрации имели такую же массу, как побеги растений при 20 мМ NaCl в сочетании с гипоксией.