Генетический анализ резистентности клещей
Генетический анализ проводят для изучения природы устойчивости и идентификации мутаций изолированных популяций. В сложных случаях определяют локализацию и группы сцепления (хромосомы) мутаций резистентности к различным препаратам.
Удобные объекты для генетического анализа — тетраниховые и фитосейидные клещи: они высокоплодовиты, имеют короткое время развития и арренотокный (гаплодиплоидный) тип размножения. Неосемененные диплоидные самки паутинных клещей откладывают гаплоидные яйца, из которых развиваются только самцы с гаплоидным (одинарным) набором хромосом. Осемененные самки откладывают как гаплоидные, так и диплоидные (оплодотворенные) яйца. Из последних развиваются самки. Фитосеииды размножаются по иному типу. У них неосемененные самки не откладывают яиц. После спаривания самки откладывают гаплоидные (будущие самцы) и диплоидные (будущие самки) яйца. Однако гаплоидные яйца образуются в результате осеменения, и на начальных этапах эмбриогенеза «самцовые» яйца диплоидны (парагаплоидия). Впоследствии же отцовский набор хромосом элиминирует (гиногенез).
Таким образом, самцы тетраниховых и фитосейидных клещей повторяют геном матери и не несут признаков отцовского генотипа. Например, если скрещивать чувствительных самок (S) с устойчивыми самцами (R), то в первом поколении (F1) все сыновья будут чувствительны и, наоборот, если скрещивать устойчивых самок (R) с чувствительными самцами (S), то самцы первого поколения (F1) будут резистентны. Анализ фенотипического выражения резистентности исходных родительских форм (К и S) и гибридов первого и второго поколений — основа генетического анализа резистентности.
Анализ гибридов первого поколения дает ответ о характере фенотипического наследования устойчивости. Для получения неосемененных самок в садки (на «плотики» или «плавающие листья») отсаживают дейтонимф паутинного клеща. Дейтонимф самок хищных клещей легко спутать с самцами. Поэтому при работе с фитосейидами в садки индивидуально рассаживают большое число нимф и после достижения ими стадии имаго отбирают девственных самок. К самкам подсаживают самцов, причем скрещивания проводят реципрокно, то есть
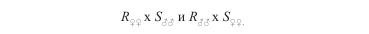
Чтобы получить максимальное число примерно одновозрастных яиц, через каждые 3 сут родительские пары пересаживают на свежие листья. После достижения потомством стадии имаго отбирают самок и обрабатывают их препаратом в серии концентраций с последующим пробит-анализом. Если пробит-линия самок первого поколения реципрокных скрещиваний совпадает с пробит-линией чувствительной популяции, то резистентность оценивается как рецессивный признак. При совпадении с пробит-линией устойчивого родителя резистентность доминанта. При промежуточном расположении пробит-линии степень рецессивности или доминантности (Д) устанавливают по формуле Стоуна:

где х1 — lgCK50 резистентной популяции; x2 — lgCK50 гетерозиготных самок первого поколения; x3 — lgCK50 чувствительной популяции.
Анализ гибридов второго поколения дает информацию о моно- или полигенном наследовании устойчивости. Для получения клещей второго поколения применяют возвратные скрещивания (беккросс) самок первого поколения с устойчивыми и чувствительными самцами. В первом случае потомство наследует фенотип родительских самок. Во втором случае происходит расщепление на два класса (RS и SS). При рецессивном наследовании устойчивости такое потомство фенотипически не будет отличаться от чувствительных родителей. Поэтому в случае рецессивного наследования анализу подвергают самцов F2, полученных от гетерозиготных самок F Самцы несут одинарный набор хромосом и непосредственно демонстрируют числовые отношения гамет самок F2. При доминантном наследовании устойчивости характер расщепления демонстрируется фенотипической реакцией смеси двух генотипов самок.
Для примера рассмотрим характер наследования резистентности хищного клеща фитосеиулюса к карбофосу. Уровень устойчивости клещей, резистентных к этому препарату, превышал чувствительность «нормальной» популяции фитосейулюса на два порядка (в 120 раз по CK50 и в 135 раз по CK95). Таким образом, разница между родительскими формами при скрещиваниях была заметной и позволяла легко диагностировать отдельные генотипы. Диапазон концентраций карбофоса, убивающих 10—95 % чувствительных клещей, лежал в пределах 0,0001—0,001 % д.в., а устойчивых — в пределах 0,01—0,25 % д.в. Клещей, полученных от скрещиваний чувствительных самок с устойчивыми самцами и устойчивых самок с чувствительными самцами, подвергали токсикологической проверке с использованием указанных концентраций карбофоса.
Устойчивость самок F1 не отличалась от родительских форм, то есть полностью наследовался доминантный признак (табл. 7, см. рис. 35).
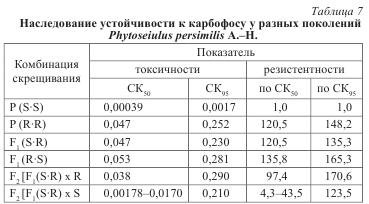
He отмечено и разницы в реципрокных (взаимно противоположных) скрещиваниях, что свидетельствует об отсутствии цитоплазматического влияния материнского организма.
Следующим этапом работы было определение числа генов, участвующих в контролировании признака резистентности. Для этого провели возвратные скрещивания гибридных самок F1 с родительскими формами.
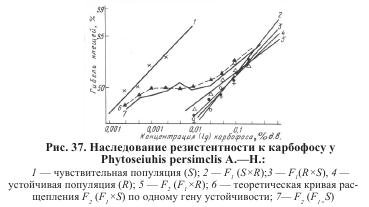
Скрещивание гибридных самок с доминантной родительской формой (резистентным самцом) F2, как и следовало ожидать, было единообразным и повторяло устойчивость резистентных самок исходной популяции (состав потомства: половина гетерозигот, половинна гомозигот по устойчивости). В анализирующем скрещивании с рецессивной родительской формой (чувствительным самцом) для диагностики потомства использовалось десять концентраций карбофоса (от 0,00025 до 0,25 %), что дало возможность обнаружить в потомстве присутствие дискретных классов клещей: одна часть особей погибла при воздействии малых доз препарата, другая — оставалась живой и при использовании высоких. Кривая смертности образовывала «плато» на уровне 50 % гибели, то есть диагностируемые классы особей составляли равные доли в потомстве.

Результаты эксперимента указывали на расщепление по одному главному гену (50 % гомозигот по чувствительности + 50 % фенотипически устойчивых гетерозигот). Поскольку величина гибели каждого из предполагаемых классов известна из токсикологических определений устойчивости отдельно родительских форм и гибридов, ожидаемую гибель вычислили как сумму гибели классов, составляющих F2, умноженную на долю каждого из классов в общем числе потомков (при расщеплении по одному гену — 0,5).
По точкам гибели, ожидаемой от каждой из использованных в опыте концентраций карбофоса, на рисунке 37 построена теоретическая кривая смертности F2. В таблице 8 приведены данные статистической проверки гипотезы методом x2:

где о — наблюдаемое число погибших клещей; е — ожидаемое число погибших; р — ожидаемое доля погибших в общем числе потомков.
Установлено, что при использовании всех десяти концентраций разница между теоретически ожидаемой и наблюдаемой гибелью клещей недостоверна, x2 < 3,8 (P>0,05), то есть гипотеза о расщеплении по одному гену, контролирующему резистентность, подтверждена.