



Клещи подотряда Acaridida
Тело длиной 300-700 мкм, сильновыпуклое или уплощенное в дорсовентральном направлении. Иногда разделено поперечной бороздкой на проподосому и гистеросому. Кожные покровы мягкие, тонкие, как правило, бесцветные или грязноватозеленые, с бронзовым, фиолетовым, розовым оттенком, у некоторых видов — тонкозернистые или с шиповатой микроструктурой. Гнатосома конусовидная. В ее состав входит пара мощных клешневидных хелицер грызущего типа, гипостом, пара ложных рутелл (жевательные лопасти) и слаборазвитые двучлениковые педипальпы. Базальные членики хелицер свободные, на внутренней поверхности неподвижного пальца клешни помимо шиповидного выроста имеется щетинка, характерная для клещей этой группы.
На спинно-боковой поверхности тела располагаются следующие группы щетинок (рис. 83, Б): на проподосоме — две пары теменных ve и vi, две пары лопаточных sсe и sci и над тазиками ног I пара латерококсальных (псевдостигмальных) органов — небольших, более или менее модифицированных щетинок характерной для данного вида формы — палочковидной, щетинковидной, перистой, ветвистой и т.д.; на гистеросоме — одна - три пары плечевых he, hi, sh, четыре пары спинных d1—d4, две-три пары боковых la, lp и одна-две пары крестцовых sae и sai. Кроме того, на гистеросоме имеется четыре пары поровидных отверстий: по одной паре вблизи плечевых и передних боковых щетинок, одна пара у заднего края тела и одна пара по обе стороны анального отверстия.
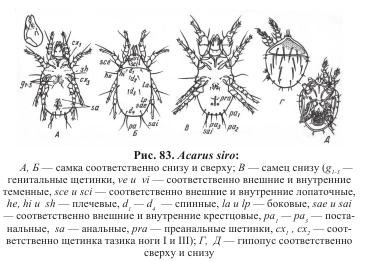
На вентральной поверхности располагается две пары тазиковых щетинок (сх1 — на тазиках ноги I и схз — на тазиках ноги III), три пары генитальных щетинок и щетинки анального комплекса: преанальные prа, анальные sa, постанальные ра и аданальные (см. рис. 83, В). У некоторых видов эти щетинки сближены между собой и образуют единую группу анальных щетинок. Количество и расположение тазиковых и генитальных щетинок весьма постоянно. Щетинки анального комплекса и спинной поверхности заметно варьируют по количеству, размеру и взаимному расположению, что служит хорошим диагностическим признаком.
Ноги состоят из шести члеников: тазика, вертлуга, бедра, колена, голени и лапки. Тазики неподвижно срослись с телом и образуют коксостернальный скелет, состоящий из эпимер, узких склеритов, ограничивающих тазики спереди, и эпимеритов — склеритов, ограничивающих тазики сзади. Ходильный аппарат включает перепончатую предлапку и непарный, большей частью когтевидный эмподий (рис. 84, А). У одних групп клещей коготок сидячий и причленяется непосредственно к вершине лапки двумя соединительными склеритами, в этом случае основание коготка уходит внутрь крупной мясистой предлапки (см. рис. 84, E). У других — коготки стебельчатые, обычно маленькие, иногда плохо заметные. С вершиной лапки они непосредственно не соединяются, а помещаются на конце более стройной и удлиненной предлапки (см. рис. 84, Д).

На ногах различают четыре типа щетинок:
- истинные — по-видимому, органы осязания (базально оканчиваются луковицей, находящейся в отверстии экзоскелета, внутренняя часть заполнена хитином, наружная часто образует различную орнаментацию — волоски, шипики, чешуйки);
- акантоиды — служат органами обоняния и вкуса (модифицированные щетинки с внутренней полостью, имеют форму гладкого шипа, расположены только в передней части лапок I и II);
- фамулюс — маленький орган конической формы, с усеченным, расширенным или даже раздвоенным дистальным концом; внутри расположен узкий канал, пересекающий корень, под ним — широкое отверстие в экзоскелете, в углублении которого находятся нервные окончания (имеется на всех стадиях развития);
- соленидии — трубки с тонкими стенками, закрытые на дистальном конце и открытые на базальном; имеют гладкую поверхность, иногда поперечно исчерчены; волосяная луковица отсутствует; корень всего очень короткий, начинается маленьким сужением (встречаются только на коленях, голенях и лапках).
Наиболее богат хетом лапки I, в состав которого обычно входит 17 щетинок (см. рис. 84, А; 85, А, Б). По расположению на лапке эти щетинки подразделяют на три группы: дистальную, медиальную и базальную. Щетинки дистальной группы размещаются вокруг основания предлапки. В состав этой группы входит девять щетинок: соленидий w3, дорсомедиальная d, дорсолатеральная наружная е, дорсолатеральная внутренняя f, вентромедиальная s, вентрола-теральные внутренние q и v, вентролатеральные наружные и и р. Вентральные щетинки s, q, v, и, р обычно короткие, шиповидные.
Медиальная группа из четырех щетинок занимает среднюю часть лапки. Сюда относятся щетинки: дорсальная bа, вентральная wa, латеральная внутренняя lа, латеральная наружная rа.
У одних групп клещей коготок сидячий и причленяется непосредственно к вершине лапки двумя соединительными склеритами, в этом случае основание коготка уходит внутрь крупной мясистой предлапки (см. рис. 84, E). У других — коготки стебельчатые, обычно маленькие, иногда плохо заметные. С вершиной лапки они непосредственно не соединяются, а помещаются на конце более стройной и удлиненной предлапки (см. рис. 84, Д).
На ногах различают четыре типа щетинок:
- истинные — по-видимому, органы осязания (базально оканчиваются луковицей, находящейся в отверстии экзоскелета, внутренняя часть заполнена хитином, наружная часто образует различную орнаментацию — волоски, шипики, чешуйки);
- акантоиды — служат органами обоняния и вкуса (модифицированные щетинки с внутренней полостью, имеют форму гладкого шипа, расположены только в передней части лапок I и II);
- фамулюс — маленький орган конической формы, с усеченным, расширенным или даже раздвоенным дистальным концом; внутри расположен узкий канал, пересекающий корень, под ним — широкое отверстие в экзоскелете, в углублении которого находятся нервные окончания (имеется на всех стадиях развития);
- соленидии — трубки с тонкими стенками, закрытые на дистальном конце и открытые на базальном; имеют гладкую поверхность, иногда поперечно исчерчены; волосяная луковица отсутствует; корень всего очень короткий, начинается маленьким сужением (встречаются только на коленях, голенях и лапках).
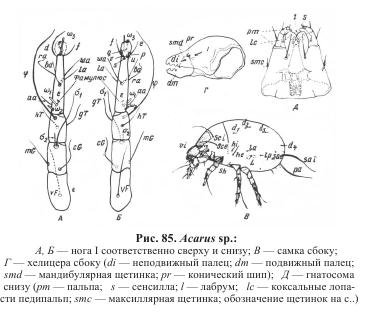
Наиболее богат хетом лапки I, в состав которого обычно входит 17 щетинок (см. рис. 84, А; 85, А, Б). По расположению на лапке эти щетинки подразделяют на три группы: дистальную, медиальную и базальную. Щетинки дистальной группы размещаются вокруг основания предлапки. В состав этой группы входит девять щетинок: соленидий w3, дорсомедиальная d, дорсолатеральная наружная е, дорсолатеральная внутренняя f, вентромедиальная s, вентрола-теральные внутренние q и v, вентролатеральные наружные и и р. Вентральные щетинки s, q, v, и, р обычно короткие, шиповидные.
Медиальная группа из четырех щетинок занимает среднюю часть лапки. Сюда относятся щетинки: дорсальная bа, вентральная wa, латеральная внутренняя lа, латеральная наружная rа.
В базальную группу входят одна тактильная щетинка аа, а также три сенсорные: два соленидия w1 и w2 и и фамулюс e. Последний расположен вблизи основания соленидия w1. Соленидий w1 появляется уже на стадии личинки, w2 — протонимфы и w3 — дейтонимфы.
Хетом лапок II—IV беднее, чем лапки I. Так, на лапках II—IV отсутствуют соленидии w2 и w3, фамулюс с, и щетинка аа, на лапках III—IV, кроме того, соленидии со и щетинки la и bа. Количество щетинок на вертлугах, бедрах, коленях и голенях ног I—IV: 1-1-4-3; 1-1-3-3; 1-0-2-2; 0-1-0-2. На голенях I—II располагается по одному длинному бичевидному соленидию ф и по две щетинки gT и hT, на голенях III и Iv — по одному соленидию ф и по одной вентральной щетинке hT. На колене I — два соленидия о1 и о2, колене II — один соленидий на обоих члениках, кроме того, по две щетинки cG и mG. На колене III — соленидий и одна вентральная щетинка mG. На бедрах I, II и IV и вертлугах I, II и III — по одной вентральной щетинке, которые обозначаются соответственно на бедрах vF.
Половой диформизм выражен в различной степени. Обычно самец меньше самки и имеет более длинные и стройные ноги. У некоторых клещей подотряда самцы бывают двух видов: гомеоморфные, близкие по строению к самке, и гетероморфные — отличающиеся более интенсивной окраской тела, длинными щетинками, гипертрофированным развитием третьей пары ног, модифицированными коготками и т. д.
Наружный половой аппарат самки состоит из генитального (яйцевыводного) и копулятивного отверстия. Первое представляет собой продольную щель и находится как правило между аподемами III и IV, второе округлое и расположено терминально. У большинства видов яйцевыводное отверстие прикрыто генитальными клапанами, смыкающимися между собой по средней линии и постепенно расходящимися назад. Под этими клапанами помещаются две пары генитальных щупалец. У многих глицифагид передний край генитального отверстия снабжен склеритом полулунной формы — эпигинием.
У самца генитальное отверстие, в котором расположен копулятивный орган, как и у самки, помещается между основаниями ног III и IV. Оно сдвинуто назад к основанию ног IV или вперед — к основанию ног I. По сторонам эдеагуса под генитальными клапанами, как и у самки, находятся две пары генитальных щупалец.
С копулятивными органами функционально связаны специальные органы фиксации. У самцов многих видов этот прикрепительный аппарат состоит из пары довольно крупных копулятивных присосок, расположенных по бокам анальной щели, и пары тарзальных присосок, сидящих на внешней поверхности ноги IV (см. рис. 83, В; 84, Г). Иногда роль прикрепительных органов играют сильно модифицированные ноги III и IV, в других случаях — предлапки ног I и II. Анальное отверстие в виде продольной щели расположено вентрально у заднего конца тела.
Предимагинальные стадии развития отличаются следующими морфологическими особенностями. Личинка имеет только три пары ног и пару коксальных органов. Последние расположены между тазиками I и представлены парой палочковидных щетинок сенсорного типа. Щетинки отсутствуют на гистеросоме ( d4, lp), на лапке I (w2 и w3) и на вертлугах I и II. Генитальное отверстие не выражено.
У протонимфы появляются четвертая пара ног, а также зачаточное генитальное отверстие и пара генитальных щупалец. Коксальные органы отсутствуют. На гистеросоме добавляются одна пара генитальных и одна пара аданальных щетинок (если последние имеются у взрослой особи), а также щетинки (d4, и lр. Для хетома ног характерно отсутствие щетинок на вертлугах I—III, четырех базальных члеников ног IV и одной сенсорной щетинки w3 на лапке I.
Гомеоморфная дейтонимфа отличается от предыдущей стадии наличием двух пар генитальных щупалец, полным набором генитальных щетинок, наличием вентральной щетинки на вертлугах I—III, щетинок w1, w2 и w3 на лапке I и полного щетинкового вооружения ног IV, характерного для взрослых особей.
Некоторые виды клещей отличаются тем, что наряду с гомеоморфной имеют гетероморфную дейтонимфу (гипопус). Он бывает подвижным или неподвижным, свободным или находится внутри протонимфальной линочной оболочки (гипопальная камера).
Неподвижный гипопус почти округлый. Гнатосома развита слабо, анальная щель короткая. Ноги сильно укороченные, так что из-под тела выступает лишь два-три членика передних ног. У некоторых видов (Acaris siro, Glycyphagus destructor) сохраняются рудименты ног и прикрепительного аппарата.
Подвижный гипопус (расселительный) овальный, с поперечной бороздкой, разделяющей тело на проподосому и гистеросому (см. рис. 83, Г, Д). Брюшная поверхность вогнутая, спинная — выпуклая. Зачаточная гнатосома имеет вид вильчатого отростка, напоминающего тритостернум гамазовых клещей, и выполняет, по-видимому, осязательные функции. На брюшной стороне, ближе к заднему краю опистосомы, расположен прикрепительный аппарат, состоящий из присасывательного диска с несколькими парами более или менее развитых присосок. Впереди прикрепительного аппарата располагаются зачатки генитального отверстия и по одной паре присосок и щетинок. С помощью прикрепительного аппарата гипопус закрепляется на поверхности насекомого-переносчика. Для этой же цели служат одна-две пары стернальных или коксальных присосок. Ноги I и II развиты сильнее, чем III и IV. Из-под тела выступают три-четыре членика, лапки вооружены многочисленными сенсорными щетинками ланцетовидной формы.
В определенных условиях температуры и влажности гипопусы линяют не на взрослых клещей, а на гомеоморфных дейтонимф, которые и дают взрослых особей — самок или самцов.
Акароидные клещи яйцекладущие. В некоторых случаях наблюдается яйцеживорождение, а иногда в теле матери последовательно развиваются все стадии — от яйца до гипопуса (Caloglypnus moniezi).
Самки спустя 1—5 сут после спаривания приступают к откладке яиц, помещая их по одному на субстрат, который служит питанием. Яйца самки откладывают в течение всей жизни (иногда до 2 мес). Оптимальные условия для развития и откладки яиц — температура и относительная влажность воздуха соответственно 20...24 °C и 80...90 %, влажность зерна — не ниже 13,8 %. Максимальная температура, при которой возможно развитие, — 30...32 °С, минимальная относительная влажность воздуха — 65 %. Плодовитость в зависимости от вида, условий питания, температуры и относительной влажности воздуха составляет в среднем около 100 яиц на одну самку.

Цикл развития акаридов представлен на рис. 86. Переход из одной подвижной стадии в другую прерывается периодом покоя и линьки. На стадии гипопуса клещи не питаются. Подвижные гипопусы мало приспособлены к неблагоприятным условиям жизни, особенно к низким температурам и низкой относительной влажности воздуха. Напротив, неподвижные гипопусы очень устойчивы, и клещи в этой стадии могут жить около двух лет, перенося кратковременные понижения температуры до -20 °C. С наступлением благоприятных условий температуры, влажности и питания из гипопусов отрождаются гомеоморфные дейтонимфы, а из последних — взрослые особи (самки или самцы). Образование гипопусов изучено недостаточно, чаще они появляются с наступлением неблагоприятных для развития условий температуры, влажности или питания. У некоторых видов (Glycypnagus domesticus, Rhizoglyphus echinopus) гипопусы обычны, у других они встречаются редко или вообще неизвестны (виды родов Tyrophagus, Gohieria).
Продолжительность цикла развития зависит от температуры, относительной влажности воздуха, условий питания (табл. 17). При благоприятных условиях акариды могут давать два-три поколения в месяц.

Амбарные клещи в большинстве очень медлительны. Расселяются они в основном с помощью ветра, а также животных. Возможно расселение и эндозоическим способом, так как гипопусы и яйца некоторых видов акароидных клещей долгое время сохраняются живыми в кишечнике животных. Подотряд Acaridida включает несколько семейств.