



Физиологические механизмы устойчивости растений к алюминию
Устойчивость к алюминию можно обеспечить с помощью двух механизмов: 1) выведением алюминия из апекса корня; 2) за счет внутриклеточной устойчивости к действию алюминия, поступающего в симпластное пространство.
Первый механизм основан на выделении карбоксилатного экссудата корневым апексом. Сведения об индуцированной алюминием экссудации органических кислот впервые были получены на корнях фасоли, причем устойчивая форма выделяла в 10 раз больше цитрата, чем чувствительная. В аналогичных условиях у пшеницы через 15 мин экспозиции выделялся малат, который был электронейтрален, поскольку сопровождался экссудацией K+. Интересно, что корни пшеницы содержали достаточно малата, чтобы поддерживать экссудацию в течение 4 ч. После 24-часовой экспозиции с алюминием (100 мкМ) было обнаружено включение 14C в малат и синтез последнего de novo. Путем скрининга 36 сортов пшеницы установлена тесная корреляция между их относительной устойчивостью к действию алюминия и количеством выделившегося малата. Однако к этим данным следует относиться с осторожностью, так как, во-первых, малат быстро разлагается почвенными микроорганизмами, и, во-вторых, его концентрация, как и способность хелатировать алюминий, достаточно низки. Растения гречихи, являющиеся аккумуляторами алюминия, напротив, секретируют оксалат с сильной алюминийсвязывающей способностью.
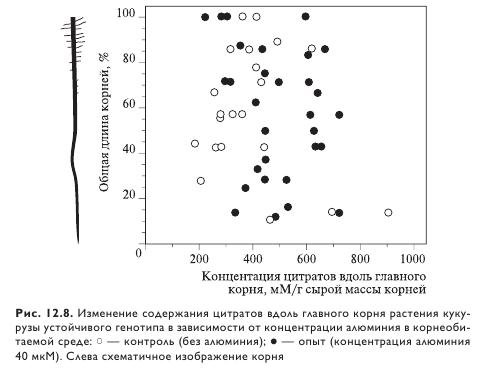
Тесная корреляция между устойчивостью к алюминию и активируемым им выделением карбоксилатов обнаружена у множества видов растений, большинство из которых — полевые культуры. В частности, синтез цитрата в ответ на воздействие алюминия отмечен у таких культур, как соя, табак, рис, сорго и кукуруза (рис. 12.8). Цитрат и малат синтезируются у рапса, овса, подсолнечника, редиса, ржи, тритикале, малат — у арабидопсиса и пшеницы, оксалат — у гречихи. Устойчивость к алюминию коррелирует с индуцируемым им выделением малата у пшеницы и арабидопсиса. Стимуляция выделения карбоксильных соединений специфична при воздействии экзогенным Al3+, причем интенсивность выделения зависит от активности алюминия в ризосфере.
Идентифицирован алюминийзависимый анионный канал корневого апекса у кукурузы и пшеницы, который может выступать в качестве транспортера, обеспечивающего выделение соответственно цитрата и малата в условиях алюминиевого стресса.
Указанные анионные каналы у пшеницы активизировались при концентрации алюминия 20—50 мкМ, при этом установлены четкие генотипические различия. Путем гибридизации кДНК изогенных линий толерантной и устойчивой к алюминию пшеницы показана большая экспрессия гена ALMT1 (от англ. Aluminium-activated Malate Transporter — активируемый алюминием транспортер малата) в рисе и культуре клеток табака. Это обеспечивало выделение малата из корней и повысило способность клеток табака к восстановлению роста после 18-часовой экспозиции в растворе, содержавшем 100 мкМ алюминия. ALMT1 может быть одним из главных генов устойчивости к алюминию. Трансгенные сорта ячменя с геном ALMT1 отличались повышенным выделением малата и более активным ростом корней при концентрациях алюминия, не превышавших 12 мкМ. Протеинкиназы, ответственные за активность анионных каналов, могут выступать в качестве посредников при выделении цитрата в ответ на алюминиевый стресс. Интересно, что концентрация цитрата или малата в корневых выделениях у устойчивых к алюминию мутантов араби-допсиса даже конститутивно (без воздействия алюминия) была выше, чем у растений дикого типа.
У ржи, обладающей среди видов полевых культур максимальной устойчивостью к алюминию, 10-часовая экспозиция в растворе, содержащем 10 мкМ алюминия, увеличивала активность цитратсинтазы на фоне усиления выделения корнями цитрата. Клеточная линия моркови также выделяла цитрат из корней и отличалась более высокой активностью митохондриальной цитратсинтазы. У сои выделение цитрата увеличивалось в течение 6-часовой экспозиции в растворе алюминия, однако только у устойчивого сорта наблюдались более продолжительная экссудация и высокая концентрация цитрата в кончиках корня. Вместе с тем активность ферментов синтеза и метаболизма цитрата в корнях сои в ответ на алюминиевый стресс изменялась незначительно. Следовательно, синтез цитрата не является ключевым звеном в выделении алюминия.
У пшеницы выделение корнем малата активируется очень быстро, но интенсивность этого процесса не повышается со временем. В этом случае алюминий, видимо, активирует уже экспрессирующийся карбоксилатный транспортер, поэтому активация генов не играет особой роли. У ржи и тритикале отмечено запаздывание активируемого алюминием выделения карбоксилата, возрастающего только через 12—24 ч алюминиевого стресса.
Повышение количества выделяемых карбоксилатов с течением времени, вероятно, обусловлено индукцией генов устойчивости к алюминию. Однако до сих пор не ясно, какая именно часть биохимического пути индуцируется. Возможны три варианта:
• увеличение количества карбоксилатных транспортеров плазматической мембраны или повышение их активности;
• возрастание интенсивности синтеза карбоксилатов благодаря увеличению количества или активности соответствующих ферментов;
• увеличение доступности карбоксилатных лигандов для транспорта.
Пока нет убедительных доказательств, подтверждающих участие какого-либо из ферментов, катализирующих в этой индуцированной ответной реакции синтез и метаболизм карбоксилатов (ФЭПК, малатдегидрогеназа, цитратсинтаза, изоцитратдегидрогеназа). Даже если под действием алюминия повышаются синтез цитрата и малата у ржи и тритикале и экссудация у устойчивых сортов, в условиях in vitro изменения активности указанных ферментов в апексах корней растений как устойчивых, так и чувствительных сортов не отмечено.
В этой связи заслуживает особого внимания карбоксилатный транспортер. У большинства видов растений алюминий быстро активирует экссудацию карбоксилатных анионов. Хотя цитоплазма содержит различные формы карбоксилатов, лишь один или два из них выделяются из цитоплазмы в ответ на действие алюминия. Активация карбоксилатного транспортера, который предположительно присутствует на плазматической мембране клеток корня, является важной частью этого механизма устойчивости. У пшеницы, как отмечалось выше, алюминий активирует выделение малата практически мгновенно, что дает основание предполагать, что именно транспорт карбоксилатов лимитирует устойчивость к алюминию.
Термодинамические условия транспорта карбоксилатов из цитозоля во внешний раствор указывают на то, что ионные каналы могут быть первичным транспортером, вовлеченным в ответную реакцию. Анионные каналы, которые специфично активируются экзогенным Al3+, идентифицированы в апексах корней у устойчивой к алюминию пшеницы и кукурузы. Активация может происходить тремя способами:
• Al3+ напрямую связывается с каналом и активирует его;
• Al3+ связывается с отдельными мембранными рецепторами, которые, в свою очередь, активируют анионный канал;
• Al3+ активирует ионный канал опосредованно через каскад сигнальной трансдукции, который может включать компоненты цитозоля.
Как показывают исследования устойчивых форм кукурузы, наиболее реальны два первых варианта.
Кроме центральной роли органических кислот, были обнаружены также другие механизмы устойчивости. Так, воздействие алюминия на устойчивые мутантные растения Arabidopsis alr-104 вызывало почти двукратное увеличение притока ионов водорода на границе раздела поверхности корня и почвы. Аналогичная тенденция отмечена и на корнях проростков кукурузы (рис. 12.9).

Повышение кислотности среды на поверхности корней вызывало выпадание в осадок алюминия из почвенного раствора, что, в свою очередь, предотвращало поступление алюминия в корни или же переводило его в менее токсичную форму. На вигне и пшенице показано, что слизь корневого чехлика связывает алюминий. Об этом свидетельствуют данные опытов, в которых удаление корневой слизи значительно увеличивало чувствительность к алюминию. Вероятно, слизь помогает сформировать диффузный барьер для алюминия или концентрирует органические кислоты, которые его хелатируют. Однако в условиях гидропоники корневая слизь у кукурузы связывала алюминий, но не защищала корни от токсического действия алюминия. Видимо, это объясняется увеличением расстояния между корневым чехликом, где секретируется слизь, и чувствительными к алюминию клетками дистальной транзитной зоны (ДТЗ).
Другой механизм устойчивости основан на хелатировании и внутриклеточной детоксикации алюминия карбоксильными анионами в симпласте уже после его проникновения в растение. Как устойчивые, так и чувствительные растения при произрастании на кислых почвах накапливают алюминий. У растений-аккумуляторов он связывается с органическими лигандами, такими, как ка-техиновые и фенольные кислоты, а также органические кислоты. Эти комплексы часто изолируются в специальных клетках, в частности в эпидермальных клетках листьев, как, например, у устойчивой к алюминию гречихи. Последняя секретирует из корней щавелевую кислоту в ответ на воздействие алюминия и накапливает нетоксичные комплексы алюминия и оксалатов в клетках листьев. В листьях гречихи алюминий присутствует в комплексе с оксалатом в соотношении 1:1, что защищает цитозоль при pH = 7,0 от повреждения. Частично устойчивость гречихи обусловлена активируемой алюминием экссудацией оксалата апексом корня. Однако гречиха, как отмечено выше, в значительных количествах аккумулирует алюминий в листьях, транспортируя его из корня в надземную часть по ксилеме в комплексе с цитратом, но не с оксалатом. Последний замещается цитратом на время транспорта по ксилеме. Затем происходит обратный обмен цитрата на оксалат, в комплексе с которым алюминий собственно и присутствует в листьях. Согласно компарт-ментационному анализу, 80% алюминия в листьях гречихи находится в вакуоли в комплексе с оксалатом в соотношении 1:3. У устойчивых гибридов кукурузы алюминий также накапливается в корнях в клеточных вакуолях.
Была предложена интересная гипотеза устойчивости к алюминию у пшеницы, основанная на ускоренной замене эпидермальных клеток корня. Во время 8-часовой экспозиции у устойчивого сорта наблюдалось точечное отмирание клеток эпидермиса корня, в то время как у чувствительного сорта такой реакции не отмечено, хотя рост корня прекращался. Видимо, замещая эпидермальные клетки, пораженные алюминием, растения устойчивого сорта поддерживают дальнейший рост корня. Следовательно, чтобы противостоять токсичному действию алюминия растения используют совокупность механизмов, при этом одни из них специфично связаны с алюминием, а другие — с более общими ответными реакциями на стресс.