



Быстрые токсические эффекты алюминия на растения
Значительное количество исследований было посвящено быстрому токсическому действию алюминия на растения. В качестве физиологического критерия токсичности использовали ингибирование роста корней, поскольку первоначальный и наиболее ранний симптом токсического действия алюминия — это быстрое (в пределах минуты) подавление роста корней. Первичной мишенью токсического действия алюминия на корень является его апекс, точнее дистальная часть проводящей зоны апекса. Сначала подавляются процессы роста клеток растяжением, а затем и делением. Увеличение экспозиции приводит к взаимодействию алюминия с ядрами клеток корня, что проявляется в повреждениях цитоскелета и нарушении деления клеток. В этих процессах велика роль фитогормонов.
Фитогормоны. Алюминий ингибирует базипетальный транспорт ИУК, что было подтверждено опытным путем. Нанесение ИУК на апикальную зону корня кукурузы, а алюминия на дистальную транзитную зону (1—2 мм от кончика корня) вызывало торможение базипетального транспорта ауксина в зону растяжения (2—5 мм от кончика корня). Интересно, что экзогенная обработка последней ИУК частично снимала индуцированное алюминием ингибирование роста корня при нанесении его на дистальную транзитную зону. Вполне возможно, что ингибирование алюминием транспорта ауксина выступает в качестве сигнала о действии данного стрессора, проходящего между зоной растяжения корня и дистальной транзитной зоной. При этом образование каллозы в плазмодесмах может быть первичным (главным) фактором в ингибировании алюминием роста корня, связанным с нарушением транспорта ауксина.
При изучении роли фитогормонов в индуцированном алюминием ингибировании роста корней пшеницы были выявлены изменения в синтезе этилена, а также в содержании и составе цитокининов в интервал времени, не превышающий 150 мин после воздействия алюминия. При выращивании проростков в водной культуре рост корней на среде с алюминием значительно (по сравнению с контролем — среда без алюминия) замедлился через 360 мин после воздействия алюминия. Выделение же этилена в отрезанных кончиках корней удвоилось после 15-минутного воздействия Al3+ и достигло максимума через 30 мин. Содержание цитокинина снизилось на 60—80% всего лишь через 5 мин воздействия алюминия, тогда как содержание зеатина увеличилось в 6 раз. Повышение содержания зеатина продолжалось и после 150-минутного периода и в 80 раз превысило уровень, зафиксированный на растениях, не подвергавшихся алюминиевому стрессу. Эти результаты доказывают, что индуцированному алюминием ингибированию роста корней предшествует значительное увеличение содержания и изменение состава цитокининов и синтез этилена. Поскольку цитокинины индуцируют образование этилена, быстрое повышение уровней цитокинина и частично зеатина, а также гидрозеатина, может способствовать ингибированию роста корня напрямую или опосредованно путем влияния на гормональный гомеостаз растения.
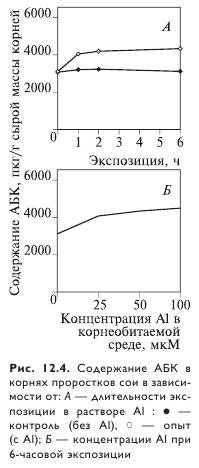
Алюминий повышал эндогенный уровень АБК в корнях сои, причем эффект зависел от экспозиции и концентрации алюминия. Концентрация АБК увеличивалась в течение первых двух часов воздействия алюминия, а затем стабилизировалась (рис. 12.4). При этом содержание АБК практически не изменялось при концентрации алюминия выше 25 мкМ. У чувствительных к алюминию сортов сои концентрация АБК в корнях в ответ на алюминиевый стресс увеличивалась сильнее, чем у устойчивых. Экзогенная обработка корней АБК повышала активность цитратсинтазы на 26%, одновременно снижая накопление алюминия в корнях на 32%. Вероятно, АБК участвует в ранней реакции корней на алюминиевый стресс.
Воздействие алюминия на пути трансдукции сигналов, выражающееся в нарушении гомеостаза внутриклеточного Ca2+ и pH, играет ключевую роль в токсичности алюминия. Алюминий может изменять уровень Ca2+ в цитозоле и его pH, а также взаимодействовать с фосфолипазой С, связанной с передачей сигналов Ca2+, и ингибировать ее действие.
В системе in vitro алюминий очень тесно взаимодействует с мембранно-связанным фосфатидилинозитол-4,5-бифосфатом (PIP2). У животных расщепление липидов плазмалеммы Р1Р2-фосфолипазой С (PLC) высвобождает инозитол-1,4,5-трифосфат (IP3) в цитоплазму. Затем IP3 путем взаимодействия с кальциевым каналом и высвобождения Ca2+ в цитоплазму формирует сигнальный каскад. В микросомальных мембранах корней пшеницы алюминий в концентрации 20 мкМ и более резко подавляет активность фосфолипазы С.
Киназы, связанные с клеточной стенкой, могут служить промежуточным звеном между клеточной стенкой и цитоскелетом цитоплазмы. Киназы пронизывают плазмалемму, причем экстраклеточная ковалентно связана с пектином клеточной стенки, а внутренняя проявляет киназную активность. Показано, что 3-часовая экспозиция корней растений арабидопсиса в растворе алюминия экспрессировала киназу, связанную с клеточной стенкой (WAK1). Это дало основание предположить, что WAK1 может участвовать в системе передачи сигнала о воздействии алюминия.
Окислительный стресс. АФК, такие, как супероксиданионы и пероксид водорода, образующиеся в процессе фотосинтеза и дыхания, могут быть вовлечены в подготовку ответных реакций на стресс. Нелетальные токсичные дозы алюминия заметно не изменяют относительные пропорции ацильных липидов в тилакоидах, а соотношение между общим содержанием галактолипидов или фосфолипидов и общим содержанием ацильных липидов остается почти постоянным. Тем не менее перекисное окисление липидов тилакоидов и образование этилена, связанные со световыми реакциями фотосинтеза, заметно активируются, что выражается в повышении скорости цепной реакции ПОЛ.
У кукурузы в условиях алюминиевого стресса наблюдается повышение концентрации фенолов, тогда как хиноновый пул хлоропластов снижается.
Одновременно ингибируется активность СОД, что может приводить к повышению уровня свободных супероксидных радикалов, активности аскорбатпероксидазы и дегидроаскорбатредуктазы, но к снижению активности глутатионредуктазы. Видимо, именно уровень активности последней in vivo является ключевым звеном пероксидазной системы хлоропластов кукурузы. Действительно, поскольку активность глутатионредуктазы ингибируется, пероксидазная система снижает свою эффективность вследствие ограничения поступления субстрата, необходимого для функционирования аскорбатпероксидазы и дегидро-аскорбатредуктазы. Кроме того, снижается активность каталазы, и, следовательно, контроль содержания H2O2, образовавшейся в процессе фотодыхания, также становится менее эффективным. Очевидно, это ингибирование связано с понижением активности СОД, что приводит к образованию гидроксильных радикалов. Однако ПОЛ усиливается лишь после длительного воздействия алюминия (24 ч и более). Таким образом, хотя активизация процессов ПОЛ не проявляется достаточно быстро, чтобы быть запускающим механизмом алюминиевой токсичности, связанное с этим образование АФК и нарушение деятельности оксидаз могут играть значимую роль в подавлении роста корней.
Цитоскелет корня. Из-за важной роли компонентов цитоскелета (микротрубочек и микрофиламентов) в процессах клеточного деления и роста корней, цитоскелет может выступать в качестве мишени алюминиевой токсичности. Алюминий может повреждать цитоскелет либо непосредственно взаимодействуя с его элементами (микротрубочками или актиновыми волокнами) или опосредованно, путем изменения каскадов передачи сигналов, например влияя на концентрацию Ca2+ в цитозоле, вовлеченного в стабилизацию цитоскелета. Отметим, что для процессов деления клетки и образования клеточной стенки важна ориентация цитоскелета. К примеру, кортикальные микротрубочки ориентируют в пространстве целлюлозные микрофибриллы, что является необходимым условием нормального роста клетки. Под действием алюминия происходит нарушение и последующая пере-стройка кортикальных микротрубочек и микрофиламентов в клетках корней. При этом значительно повышается натяжение актиновых филаментов клеток, показанное, в частности, для сои. Индуцируемые алюминием структурные изменения клеток, видимо, лежат в основе морфологических изменений и структурных дефектов корней, подвергшихся алюминиевому стрессу.
Образование каллозы и отложение лигнина. Каплоза представляет собой полисахарид, состоящий из цепей 1,3-в-глюкана и образующегося на определенном этапе формирования клетки в ответ на повреждение. Образование каллозы в корнях является ранним симптомом токсичности алюминия. Методом флуоресцентной спектрометрии каллозу можно обнаружить в кончиках корней сои уже через 2 ч после начала воздействия алюминия в концентрации 50 мкМ, а у чувствительных к алюминию сортов пшеницы — при 75 мкМ. На основании этих и других данных можно было бы даже предложить быстрый и чувствительный метод обнаружения фитотоксичного эффекта алюминия, основанный на синтезе каллозы. Однако синтез каллозы не обнаружен у двух чувствительных к алюминию мутантов арабидопсиса, что указывает на отсутствие прямой связи между индуцированным алюминием ингибированием роста корней и отложением каллозы.
Лигнины, как ароматические компоненты, представляют собой неотъемлемую часть вторичной клеточной стенки. Отложение лигнинов в ответ на воздействие алюминия было обнаружено в коровых клетках, расположенных на расстоянии 1,4-4,5 мм от кончика корня (зона растяжения) пшеницы после 3-часовой экспозиции в растворе алюминия с концентрацией 50 мкМ. При этом лигнин находился в клетках с поврежденными в результате действия алюминия мембранами, что дало основание рассматривать лигнификацию в качестве маркера токсичности алюминия, поскольку она коррелировала с торможением процессов удлинения корня. Важно подчеркнуть, что кДНК у пшеницы, индуцированная алюминием, демонстрировала высокую гомологию с геном фенилаланин-аммонийлиазы — ключевого фермента биосинтеза лигнина.