



Клеточный уровень адаптации растений к засолению
В литературе нет данных, указывающих на адаптацию ферментов к засолению. Поэтому механизмы адаптации на клеточном уровне заключаются в том, чтобы воспрепятствовать концентрированию солей в цитоплазме путем их компартментации в вакуоли. То, что это имеет место у большинства видов, потверждается концентрированием значительных количеств соли в листьях (свыше 200 мМ), которые при этом нормально фотосинтезируют. Вместе с тем подобные концентрации in vitro полностью подавляют активность ферментов.
В общем, Na+, как и Cl-, начинает ингибировать активность большинства ферментов при концентрациях выше 100 мМ. Если Na+ и Cl- задерживаются в вакуоли, органические растворы преимущественно в виде пролина и глицинбетаина должны накапливаться в цитоплазме и органеллах, чтобы сбалансировать осмотическое давление ионов в вакуоли.
Внутриклеточная компартментация и синтез осмопротекторов. Самый эффективный путь поддержания низкой концентрации Na+B цитоплазме — удерживание его в вакуоли каждой клетки. Чтобы сохранить равные осмотические потенциалы в вакуоли и цитоплазме в последней должны накапливаться совместимые растворы, не оказывающие негативного влияния на метаболизм клетки. Важную роль при этом играет большой объем вакуоли, где сохраняется Na+.
Накопление натрия в вакуоли. В процессе внутриклеточной компартментации Na+ поступает в клетки листа и закачивается в вакуоль, прежде чем содержание Na+ в цитоплазме увеличится до значимого уровня. Этот процесс катализируется вакуолярным Na+/H+-aнтипортером. Концентрационный градиент H+ первоначально устанавливается благодаря Н+-АТФазе и пирофосфатазным белкам. Активность №+/Н+-антипортера увеличивается при добавлении Na+, что показано на корнях ячменя, томатов, подсолнечника, причем это усиление заметнее у солеустойчивых видов по сравнению с солечувствительными. Последнее особенно наглядно показано для риса. Интересно, что индукция сказывается на повышении уровней транскриптов у некоторых членов семейства генов AtNHX у арабидопсиса, кодирующих вакуолярные Тф+/Н+-антипортеры. Засоление индуцирует также активность вакуолярных Н+-насосов, что характерно как для солеустойчивых, так и солечувствительных видов.
Особого внимания заслуживают ионные каналы и переносчики, регулирующие движение солей через мембрану (см. рис. 10.2). Специфический переносчик Na+ отсутствует. Поступление Na+ в клетку осуществляется на фоне конкуренции с другими катионами, в особенности с K+, с помощью переносчиков, обладающих высоким сродством к K+, а также через неселективные Са2+-зависимые катионные каналы. Через последние могут поступать большие количества Na+ из сильно засоленных почв, особенно в случае отсутствия селективного контроля за этим процессом. Натрий может выводиться из цитоплазмы с помощью Na+/Н+-антипортеров, управляемых трансмембранными градиентами pH.
Эти транспортные процессы функционируют совместно и согласованно, контролируя нетто-поглощение Na+ клеткой. Внутриклеточная компартментация Na+ происходит при участии вакуолярного Na+/H+-aнтипортера, управляемого градиентом pH на тонопласте. Природа транспортеров, поддерживающих низкую концентрацию Na+ в органеллах, таких, как хлоропласт и митохондрий, до сих пор неизвестна. Что касается транспорта Cl”, то у некоторых видов он связан с солеустойчивостью.
Концентрация Na+ в вакуоли действительно может играть одну из ключевых ролей в солеустойчивости у целого ряда видов. Так, сверхэкспрессия вакуолярного Na/Н+-антипортера (NHX1) у арабидопсиса повышает солеустойчивость арабидопсиса, томатов и рапса, а сверхэкспрессия гена нативной вакуолярной Н+-транспортирующей пирофосфатазы увеличила солеустойчивость арабидопсиса. Важно, что при этом не нарушаются pH цитоплазмы и вакуоли, а также баланс калия.
Механизм, лежащий в основе повышения солеустойчивости в результате действия вышеуказанных переносчиков, пока не ясен. Однако совершенно очевидно, что важно не просто усиленное выведение из побега Na+ (по крайней мере у арабидопсиса), а способность эффективно компартментировать его.
Этот вывод очень важен в силу нескольких обстоятельств. Во-первых, удивительно, что сверхэкспрессия одного нативного гена может вызвать такое резкое увеличение солеустойчивости. К сожалению, этот подход до сих пор не был реализован в селекции. Вполне возможно, что в этом случае повышение солеустойчивости будет достигнуто за счет потери урожайности, что необходимо проверить опытным путем. Во-вторых, повышение активности компоненты тонопластного транспорта может привести к плейотропному повышению активности других генов или ген-продуктов для обеспечения соответствующих уровней накопления Na+. Так, для сверхэкспрессии активности пирофосфатазы, транслоцирующей H+ (может в свою очередь вызвать избыточное накопление Na+ в вакуолях), необходимо поддерживать повышенную активность вакуолярного Na+/Н+-антипортера и высокую скорость синтеза осмопротекторов. Механизмы этих процессов на сегодня неизвестны. В частности неясно, происходит ли ускоренная трансляция или транскрипция необходимых генов или повышение активности всех процессов протекает на посттрансляционном уровне. Следует заметить, что конститутивная сверхэкспрессия протеинкиназы также увеличивает как солеустойчивость, так и накопление Na+ у арабидопсиса.
Утечка натрия из вакуоли. Одновременно с закачкой Na+ в вакуоль происходит его утечка в цитоплазму (вероятно через ионные каналы). Катионселективные каналы, так называемые SV- и FV-каналы, присутствуют в вакуолярной мембране растительной клетки и проницаемы как для Na+, так и для K+. Если они в интакт-ной клетке открыты, это может вызвать проблемы в удержании Na+ в вакуоли. Однако активность этих каналов достаточно низкая и, как показано, они не различаются у видов Plantago с контрастной солеустойчивостью. Интересно, что в этом опыте активность каналов у растений, выращенных при сильном засолении, значительно ниже, чем при слабом засолении, что указывает на возможные значительные энергетические затраты по закачке вышедшего в цитоплазму Na+ обратно в вакуоль. Таким образом, на сегодня мы мало знаем о контроле утечки Na+ из вакуоли и, следовательно, о возможных манипуляциях по увеличению способности клеток удерживать Na+.
Синтез осмопротекторов. По мере накачивания Na+ в вакуоль осмотический потенциал последнего должен быть сбалансирован с осмотическим потенциалом цитоплазмы. Это достигается путем синтеза и накопления в цитоплазме органических растворов, не ингибирующих биохимические реакции. Более того, они защищают их от повышенных концентраций неорганических ионов. Эти метаболиты нейтральны, высокорастворимы и включают сахарозу, пролин, глицин-бетаин, манит (рис. 10.10).

Созданы трансгенные формы растений, отличающиеся сверхвысоким накоплением этих растворов благодаря стимулированию синтеза, ингибированию распада или ускоренному транспорту к растущим тканям. За исключением пролина трансгенные манипуляции не приводят к осмотически значимому накоплению метаболитов. Смысл большинства действий на сегодня — это не просто сбалансировать осмотический потенциал в цитоплазме и таким образом содействовать большему накоплению соли в вакуоли, а защищать растение от повреждения, помогая поддерживать структуру белка или увеличивая устойчивость к АФК. Расчет метаболических затрат подобных манипуляций возможен, однако для минимизации отрицательного эффекта превращения фотоассимилятов в эти растворы их синтез должен быть индуцибельным в результате действия засоления. Разделение воздействия этих компонент на соле- и засухоустойчивость представляет трудности, особенно ввиду того, что модельным объектом, на котором впервые было показано явление сверхаккумулирования, служил табак, более чувствительный к осмотическому компоненту NaCl, чем к самим ионам. Тем не менее у некоторых растений, например кукурузы, наблюдается тесная корреляция между концентрацией осмопротекторов и солевыносливостью. Видимо, эти компоненты достаточно эффективны как в поддержании отрицательного осмотического потенциала в цитоплазме, так и в прямой защите белковых и рибосомальных структур от отрицательного воздействия повышенных концентраций Na+ в цитоплазме.
Хотя осмопротекторы и помогают растению в некоторой мере переносить засоление, тем не менее нужно компартментировать значительное количество Na+, поэтому активный синтез осмопротекторов должен сочетаться с плейотропным повышением активности натриевого насоса.
Устойчивость к высокой концентрации Na+ в цитоплазме. Активность ферментов и рибосомальная активность устойчива к Na+ в цитоплазме до достаточно высоких его концентраций (около 100 мМ) в присутствии осмопротекторов и даже метаболических интермедиатов типа фосфоенолпирувата (ФЕП). Однако, поскольку чувствительность к солям, как ферментов, так и рибосом, сильно зависит от малейших изменений в условиях определения активности ферментов в системе in vitro, полученные выше данные могут и не отражать происходящее in vivo. Только одновременные определения in vitro и in vivo в сочетании с мониторингом содержания субстратов и Na+ могут дать относительно реальную картину.
Вместе с тем у галофитных бактерий, у которых изменили структуру некоторых ферментов для достижения устойчивости к высокой концентрации Na+ (благодаря большему числу кислотных остатков на поверхности белков-ферментов) для проявления ферментативной активности требовалась высокая концентрация Na+. Поэтому генная инженерия солеустойчивых ферментов в растениях может потребовать изменений не только в структуре белков, но и снижения метаболической эффективности при выращивании на незасоленных почвах. Изменение концентраций осмопротекторов, помогающих поддерживать структуру белка при повышенной концентрации Na+, представляется более реальным решением проблемы.
Реакции на повреждение и репарация. В ответ на солевой стресс синтезируется целый ряд белков. Многие из них, такие, как осмотины и дегидрины, обладают свойствами, близкими к шаперонам и, видимо, вовлечены в процессы поддержания структуры белка при сильном засолении и в тех случаях, когда взаимодействие белков с водой в силу тех или иных обстоятельств нарушено. Благодаря высокой гидрофильности они могут также стабилизировать мембранную структуру. Одни из этих стресс-белков проявляют свои действия в ответ на неожиданный осмотический шок экспериментального воздействия, другие вовлечены главным образом в реакции растений на осмотический компонент солевого стресса. Обнаружение специфичных стресс-белков ответа на высокую концентрацию Na+ требует соответствующего осмотического контроля. Их можно выявить по крайней мере в побегах, благодаря относительно позднему появлению (дни-недели) после добавления NaCl.
Конститутивная сверхэкспрессия белков позднего эмбриогенеза (LEA) ячменя в рисе определяла солеустойчивость последнего, хотя сходный эффект после добавления маннита предполагает, что LEA защищает скорее от осмотического эффекта NaCl, нежели от ионспецифичного компонента. Конститутивная сверхэкспрессия в табаке БТШ солеустойчивой бактерии повышает солеустойчивость при одновременном снижении содержания Na+ в побеге и не оказывает никакого влияния на растение в бессолевом варианте. Осмопротектор глицин-бетаин также выступает как шаперон и снижает ПОЛ, а также защищает митохондриальный транспорт электронов. Синтез полиаминов (путресцина и спермина) усиливается в ответ на стресс, в том числе солевой, а увеличение содержания путресцина заметнее у солевыносливого, чем у солечувствительного томата. Функция полиаминов — защита главным образом от свободных радикалов.
Снижение образования АФК (рис. 10.11) в результате ингибирования фотосинтеза и/или усиление их тушения представляется важной компонентой реакции растений на засоление и другие стрессы.
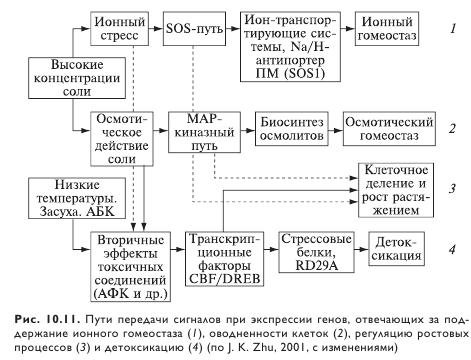
Интересно, что активность ферментов, вовлеченных в удаление активного кислорода, выше у дикого солеустойчивого томата, чем у его близкородственного культурного сорта. Это предполагает, что удаление АФК представляет собой скорее часть механизмов защиты против засоления у этого вида, нежели ответ на солеиндуцированное повреждение, опосредованное осмотическим компонентом солевого стресса. Хотя разумнее снизить повреждение, чем обеспечивать репарационные процессы, один из немногих устойчивых к высокой концентрации Na+ мутантов обладает этим свойством благодаря мутации ферментов, причастных к удалению АФК. Несмотря на то что повышенные уровни АФК могут быть результатом прежде всего осмотического стресса, вызывающего снижение устьичной проводимости, тот факт, что мутация обеспечила определенную защиту против умеренных концентраций LiCl, предполагает наличие значительного ионспецифичного компонента у этого эффекта. Идентификация солечувствительного мутанта путем скрининга на чувствительность к ультрафиолету указывает на то, что определенные звенья (элементы) солевыносливости могут быть связаны с процессом репарации ДНК.