



Адаптация к засолению на уровне целого растения
Солеустойчивость зависит от способности растения контролировать транспорт соли в ходе нескольких процессов.
Избирательность поглощения клетками корня. До сих пор не ясно, какие типы клеток управляют избирательностью поглощения ионов из почвенного раствора. Первоначальное поглощение Na+ и Cl- может происходить в эпидерме, экзодерме или, если почвенный раствор транспортируется по апопласту, даже в эндодерме.
Загрузка ксилемы. Имеются доказательства преимущественной загрузки K+ (а не Na+) клетками корневой стели.
Удаление соли из ксилемы в верхней части корней, стебля, черешка или влагалища листьев. У многих видов Na+ задерживается в верхней части корневой системы и нижней части стебля, что указывает на обмен K+ на Na+ в клетках стели корня или в сосудистых пучках стебля и черешков.
Загрузка флоэмы. Транспорт Na+ или Cl- по флоэме незначителен (особенно у более солеустойчивых видов), что указывает на отсутствие экспорта солей к растущим частям побега.
Выведения через солевые железы. Специализированные типы клеток имеют лишь галофиты, которые обладают всеми механизмами контроля поглощения, транспорта и выделения солей. Гликофиты обладают лишь двумя первыми механизмами и реализуют их в разной степени. Генетические вариации в рамках конкретного вида или между близкородственными видами обусловлены преимущественно различиями в контроле за поглощением солей корнями или загрузкой ксилемы. В частности, генетические вариации в загрузке Na+ в ксилему объясняют различия в накоплении Na+ и соответственно в солеустойчивости у видов рода Triticum. Выведение солей особенно важно для многолетников, листья которых живут год или более. В этом случае возникает острая необходимость регулировать поступающий солевой поток в течение гораздо большего периода времени, чем у однолетников, листья которых живут около одного месяца. Поддержанию низкой скорости накопления соли в листьях способствуют также высокое соотношение побег/ корень, высокая скорость роста, отсутствие апопластного транспорта солей в корнях.
Регуляция транспорта Na+ в побег. Механизмы, способствующие выживанию растений в условиях сильного почвенного засоления, включают поддержание низкой концентрации Na+ в побегах. Последнее отрицательно коррелирует со скоростью роста. Сорта полевых культур, устойчивые к засолению, отличаются меньшей концентрацией солей в побегах. Однако эта закономерность не всегда действует при сравнении разных видов. Например, ячмень и хлопчатник относятся к относительно солевыносливым видам и переносят более высокие концентрации Na+ в побегах, чем пшеница.
Поддержание низкой концентрации Na+ в побегах сочетается с высокой концентрацией K+. В действительности, более высокое соотношение K+/Na+ представляется для многих видов более важным, чем просто поддержание низкой концентрации Na+. Последнее имеет смысл при условии, что натриевая токсичность преимущественно обусловлена конкуренцией с K+ за К+-связывающие сайты. В связи с этим интересно подчеркнуть, что SOS1-, SOS2- и SOS3-мутанты имеют дефекты в калийном питании, а также в транспорте Na+.
Пути поступления в корень. Первоначальное поступление Na+ из почвенного раствора в коровую паренхиму осуществляется пассивно за счет градиента концентрации и потенциала. Нетто-накопление Na+ в растении равно разнице между пассивным поступлением и активным выходом. Выведение Na+ из корня в зоне первоначального его поступления представляет собой важный механизм уменьшения повреждения некоторых растений на засоленной почве.
Скорость поступления Na+ в клетки корня составляет 0,5—2 мкМ/г сырой массы в минуту для заметно отличающихся друг от друга культур (пшеница, рис, арабидопсис) при внешней концентрации Na+ — 50 мМ. Подобная, достаточно высокая скорость поступления не приводит к быстрому накоплению Na+. Концентрация Na+ в корне с течением времени остается достаточно постоянной, а в побеге имеет тенденцию к медленному повышению. Интересно, что скорость поступления Na+ у галофитов заметно ниже, чем у гликофитов, по крайней мере, при концентрациях NaCl не превышающих 100 мМ. Ситуация может измениться при концентрациях NaCl, токсичных для гликофитов.
Возможны три пути поступления Na+ в растения. Два из них опосредованы белками и отличаются чувствительностью к добавлению внеклеточного Ca2+. Третий путь возможно обусловлен «утечкой» Na+ в корень через апопласт. Относительный вклад каждого из этих путей в общее поступление Na+ колеблется в зависимости от вида и условий выращивания. Хотя вероятнее всего, что каждый их этих путей структурно независим, каждая структура может реализовать более одного пути поступления Na+. Так, частичная чувствительность неселективного катионного канала к Ca2+ в корнях пшеницы указывает на возможную причастность этих каналов как к Са2+-чувствительному, так и к Са2+-нечувствительному компоненту поступления Na+.
Ca2+-чувствительный путь. Известно, что токсичный эффект Na+ можно ослабить добавлением в солевой раствор до 10 мМ Ca2+. Хотя влияние Ca2+ комплексное, защитный эффект кальция против действия Na+ обусловлен по меньшей мере частичным ингибированием накопления Na+B корнях и побегах растения, а также стимулированием накопления K+. Отмеченное пониженное накопление Na+ объясняется в том числе некоторым ингибированием кальцием однонаправленного поступления Na+ в корни. Одновременно Ca2+ снижает стимулированный натрием выход K+. При рассмотрении взаимодействия Na+ и Ca2+ нельзя игнорировать влияние NaCl на активность Ca2+. Более того, видимый конкурентный эффект NaCl на связывание кальция с белками, клеточными мембранами и стенками или на транспорт и питание кальцием необходимо отделять от простого снижения активности Ca2+ в растворах с высокой ионной силой.
Согласно одной из гипотез, влияние внеклеточного Ca2+ на транспорт Na+ и K+ можно отнести на счет SOS-сигнального пути (рис. 10.8). Согласно этой модели, засоление вызывает повышение концентрации Ca2+ в цитозоле, активирующее SOS3, что приводит к изменению экспрессии и активности Na+- и К+-транспортеров. Степень повышения концентрации цитозольного Ca2+ зависит от его внеклеточной концентрации. SOSS-мутанты экспрессируют мутантную форму SOSS-белка, которая, как предполагается, менее чувствительна к Ca2+, поэтому SOS3-Myтантам необходима более высокая концентрация внеклеточного Ca2+ для роста корней при засолении, чем дикому типу арабидопсиса. Однако данную гипотезу необходимо экспериментально проверить. Вместе с тем на пшенице показано, что внеклеточный Ca2+ ингибирует однонаправленное поступление Na+, а также поступление Na+ через неселективный катионный канал. Видимо, эффект Ca2+ на поступление Na+ может быть прямым, без сигналов для модификации активности ионных каналов, исходящих из цитоплазмы. Сходный эффект внеклеточного Mg2+ на поступление Na+ укрепляет последнюю гипотезу.
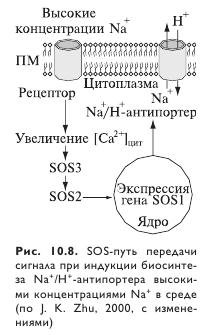
Имеется много доводов в пользу того, что именно неселективные катионные каналы являются наиболее вероятным путем для чувствительного к Ca2+ поступления Na+. Несмотря на наличие многих генов-кандидатов, которые могут кодировать эти каналы, их точная молекулярная идентичность остается невыясненной. Учитывая сложную гетерогенную структуру корня, можно лишь предположить существование нескольких путей для Са2+-чувствительного поступления Na+, кодируемых несколькими генами.
Ca2+-нечувствительный путь. Нечувствительный к Ca2+ компонент поступления Na+ может быть обусловлен, по меньшей мере частично, его поступлением через неселективный катионный канал (NSCC). Однако возможны также другие пути, кодируемые представителями семейства генов НКТ, KUP и НАК.
HKT1-транспортер из пшеницы, экспрессированный в дрожжах, катализирует высокоаффинный На+/К+-симпорт, а при повышенных концентрациях Na+ может катализировать низкоаффинный Na+-yнипорт с низким сродством. Этот тип поступления Na+, видимо, нечувствителен к Ca2+. Однако мембраны, на которые нацелены действия HKT1 в интактных растениях, пока не известны. По некоторым данным этот ген все-таки вовлечен в поступление Na+. Остается неясным, имеет ли Na+-транспортирующая способность HKTl отношение к поступлению Na+ через плазмалемму клеток интактного корня.
Обходной путь (шунт). Третий путь поступления Na+ в растения, видимо, обусловлен его поступлением в корень через апопласт. Добавление Ca2+ оказывает лишь незначительный эффект на солевыносливость сортов риса и поглощение Na+. Вместе с тем растения риса с высокой концентрацией Na+ в побеге обладали большим током воды через апопласт. Вклад апопластного пути в общее поступление Na+ в растение сильно зависит от вида, так для риса он был в 10 раз больше, чем для пшеницы.
В этой связи интересны наблюдения за анатомической адаптацией галофитов, направленных на ограничение апопластного поступления соли в растение. Ширина поясков Каспари у галофитов в 2—3 раза больше, чем у негалофитов, а внутренний слой коровых клеток может дифференцироваться во вторую эндодерму.
Выход Na+ из корня. Нетто-аккумуляция Na+ в коровых клетках корня является результатом двух процессов: 1) поступления через ионные каналы; 2) выхода благодаря №+/Н+-антипортеру, хотя последнее еще не доказано (рис. 10.9).
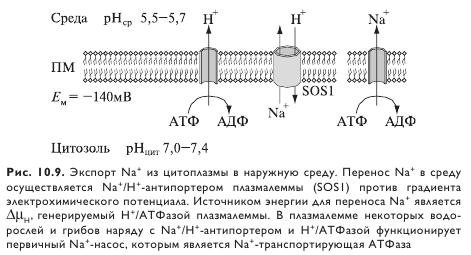
Уменьшение накопления Na+ и, как следствие, повышение солевыносливости может быть обусловлено или снижением поступления, или повышением выхода Na+. Важный вопрос в отношении Na+/Н+-антипортера — стехиометрия обмена. Электронейтральный антипортер запускается при засолении, если значение pH вне клетки увеличивается в сторону электронейтральности. В общем, именно стехиометрия определяет метаболические затраты по вытеснению Na+. При обмене Na+ на H+ в соотношении 1:1 и, соответственно, такой же стехиометрии затраты составят 1 моль АТФ на один вытесненный ион Na+ (при условии, что на вытеснение 1 протона АТФазой плазматической мембраны требуется гидролиз 1 моль АТФ). Однако теоретическая термодинамическая стоимость вытеснения Na+, т. е. энергия, необходимая для переноса 1 иона Na+ из цитозоля в наружный раствор против электрохимического градиента, видимо, намного меньше энергии, требующейся для обмена Na+ на H+. Она зависит от уровня засоления, скорости поступления соли и внутриклеточного транспорта. Таким образом, корни могут брать на себя неоправданно высокие затраты энергии по вытеснению Na+.
Регуляция нетто-поглощения Na+. Скорости однонаправленного поступления Na+, измеренные у гликофитов, могут превышать нетто-поглощение на порядок или более, что подразумевает высокие скорости его выхода. Остается неясным, почему так высока скорость поглощения и что определяет скорость нетто-поглощения?
Высокая скорость однонаправленного поступления натрия типична для гликофитов и определяется с помощью механизмов, являющихся преимущественно не №+-селективными. Транспортеры, задействованные в процессе поступления натрия, вероятно, участвуют в поглощении и других катионов, например NH4+ или Ca2+. Нормальная физиологическая роль этих путей поступления Na+ остается неизвестной, как и то, можно ли изменить селективность и активность этих механизмов с целью снижения транспорта Na+ без ущерба для других функций. Видимо, у гликофитов нетто-поглощение натрия в меньшей мере регулируется через контроль поглощения, чем через выход избыточного количества Na+ с помощью Na+-селективных Na+/Н+-антипортеров. В связи с этим возникает естественный вопрос — почему поступление натрия не избирательное, как выход, и актуален ли такой механизм для галофитов, выращиваемых при засолении?
Высокие скорости как поступления натрия в корни, так и выхода из них ставят и другой интересный вопрос, например, как корни определяют (измеряют) скорость нетто-поступления натрия? Как отмечалось выше, концентрация Na+ в корнях менее вариабельна, чем в побегах, и часто колеблется лишь незначительно среди видов или сортов, сильно отличающихся накоплением Na+ в побегах (рис, ячмень, кукуруза, пшеница и др.). Более того, концентрация Na+ в корнях определяется внешней концентрацией NaCl и у некоторых видов повышается при умеренном засолении. Это указывает на наличие некой критической точки в корнях, регулирующей концентрацию Na+ и, возможно, даже его нетто-поступление. Клетки корня каким-то образом чувствуют внутриклеточный уровень натрия и соответственно контролируют транспортеры натрия. В отличие от побегов корни могут контролировать скорости выхода натрия в двух направлениях — из корня в наружный раствор и в ксилему для транспортировки в побег. Вполне возможно, что экспорт в побег может происходить как побочный эффект процессов в корне, поддерживающий его гомеостаз. Так, у пшеницы ингибирование однонаправленного поступления натрия путем добавления Ca2+ оказывает больший эффект на концентрацию Ca2+ в побегах, чем в корнях. Видимо, концентрация Na+ в корнях контролируется независимо от скорости поступления, в то время как транспорт натрия в побег зависит от скорости выхода из корня избыточного количества Na+. Таким образом, хотя снижение поступления Na+ может оказывать слабое влияние на концентрацию Na+ в корне, оно может сильно снизить накопление натрия в побегах.
Высокие скорости как поступления Na+, так и его выхода подразумевают, что даже маленькие изменения каждого из них вызовут существенные изменения в нетто-поглощении. Предположим, что два растения имеют одинаковую скорость поглощения, а скорость выхода Na+ составляет 90 и 95% от скорости поглощения. При этом скорость нетто-поглощения первого будет в 2 раза (10:5) выше, чем у второго, хотя различия между ними по скорости транспорта натрия мизерные. Более того, если эти растения будут поддерживать те же концентрации Na+ в корнях, поглощение натрия побегом будут различаться более чем в 2 раза. Таким образом, даже очень маленькие различия в активности транспортера могут привести к существенным различиям в накоплении Na+ в побегах.
Контроль за загрузкой ксилемы. Чтобы избежать накопления избыточного количества Na+ в побегах путем поддержания его низких концентраций в ксилеме, необходимо снизить его поступление в ксилему из симпласта корня или увеличить его выведение из ксилемы, пока он не достиг чувствительных тканей побега. Для управления транспортом Na+ по растению требуются особые типы клеток, локализованные в совершенно конкретных зонах растения и катализирующие транспорт согласованно. Так, для сокращения поступления Na+ в побег в апопластном компартменте ксилемы клеткам внешней части корня необходимо снизить поступление натрия из почвенного раствора и/или увеличить его выход в почвенный раствор. Одновременно клетки внутренней части корня должны увеличить поступление из ксилемного раствора и/или снизить поступление в ксилемный раствор. Таким образом, чтобы регулировать поступление натрия в побег, нужно манипулировать транспортными процессами в плазмалемме во внутренней и наружной частях корней, т. е. с каждой стороны апопластного барьера — эндодерме и экзодерме в противоположных направлениях. Для углубления понимания сложных процессов адаптации к засолению на уровне целого растения требуется больше данных о последствиях манипуляций с транспортерами и сигнальными элементами в специфичных видах клеток.
Рассматривая энергетику передвижения Na+ в ксилему из паренхимных клеток стели, можно предположить, что это передвижение активное. Различия в активности Na+ достаточно малы. Концентрация Na+ в ксилемном соке находится в диапазоне 1—10 мМ. Сходные величины (1—30 мМ) обнаружены в цитоплазме клеток корня. В таком случае различия в энергии будут определяться главным образом разностью потенциалов между двумя сторонами мембраны, которая равна примерно 100 мВ и отрицательна внутри паренхимных клеток по отношению к ксилемному соку. Тогда поступление Na+ в ксилему будет активное, т. е. накопление Na+ в побегах управляется насосом. Таким образом, накопление Na+ в побегах управляется его активной закачкой в ксилему корня, что показано на арабидопсисе при умеренном засолении. Происходит ли то же самое у других культур, в особенности у зерновых, остается неясным.
Возможно также, что транспорт Na+ в ксилему происходит пассивно, по крайней мере у сильно транспирирующих растений. Если же концентрация Na+ в клетках стели выше, чем говорилось ранее (около 100 мМ), а в ксилеме — у нижней границы упомянутого выше диапазона (около 2 мМ), что было показано на декапитированных растениях ячменя в условиях, лимитирующих интенсивную транспирацию, то разница в энергии может способствовать пассивному поступлению Na+ в ксилему.
Это обсуждение показывает, как мало известно о транспорте Na+ в ксилему и энергетике этого процесса. Возможно, что рассмотренная выше схема процесса неверна, и загрузка натрия активна при низкой засоленности и пассивна при высокой.
Уместно напомнить также о роли АБК в загрузке ксилемы. АБК ингибирует выход K+ и Cl- в ксилему, однако о контроле АБК загрузки Na+ в ксилему известно мало. Вместе с тем добавление АБК стимулирует вытеснение H+ в ксилему, что, в свою очередь, может скорее стимулировать, чем ингибировать любую активность Na+/H+-антипортеров в плазматической мембране паренхимных клеток ксилемы.
Другой потенциальный регулятор загрузки Na+ — инозитол стимулирует перенос Na+ в побег у солевыносливой хрустальной травки. В интактных растениях инозитол синтезируется в листьях в ответ на засоление и транспортируется к корням по флоэме. Предполагается, что именно это служит сигналом на дальнем расстоянии о водном стрессе в побегах, стимулируя поглощение Na+, которое, в свою очередь, снижает осмотический потенциал побега.
Идентифицированы мутанты арабидопсиса, накапливающие высокие концентрации Na+. В одном из них (SOS1) накопление Na+ в побеге увеличилось в 2—7 раз, а в корнях практически не изменилось, что, по мнению исследователей, обусловлено сниженным контролем за загрузкой Na+ в ксилему. Выявление молекулярной основы мутации позволит лучше понять причину указанного феномена.
Выход натрия из ксилемы. Другим путем снижения накопления Na+ в побеге могло бы быть его выведение из ксилемы до того, как он достигнет молодой части побега. Это может происходить: а) в зрелом корне; б) в мезокотиле; в) в основании побега; г) в зрелой части побега. Точные механизмы данного процесса пока не ясны, но один из наиболее вероятных путей — специальный канал, обеспечивающий проницаемость внутрь для Na+ и уже описанный для паренхимных клеток ячменя. Возможен и другой путь, согласно которому при сильном засолении Na+/Н+-антипортер работает в обратном направлении, т. е. закачивает Na+ в цитозоль. Однако при условии примерно 100-кратной разницы в активности H+ между цитоплазмой и апопластом (если обмен элетронейтрален) предложенный антипортер в любой физиологически приемлемой ситуации вряд ли термодинамически возможен. Действительно, тогда концентрация апопластного Na+ должна быть в 100 раз выше, чем обнаруженная в цитоплазме (около 10 мМ). Тем не менее возможно, что Na+/Н+-антипортер не всегда действует in vivo как электронейтральный 1Na+/1Н+-антипортер.
Интересно, что в условиях аноксии корней кукурузы имело место увеличение переноса Na+ из корней в побег при слабом засолении (11 мМ) и ингибирование переноса при умеренном засолении (40 мМ). Это согласуется с механизмом активного выведения Na+ из ксилемы при относительно слабом засолении и пассивного — при более сильном засолении. При этом необходимо иметь в виду влияние деполяризации мембраны на пассивное поступление Na+ и загрузку ксилемы.
Весьма заметной при этом может быть роль передаточных клеток. В корнях похожие на передаточные клетки протуберанцы можно видеть вблизи ксилемы, в проксимальной части корня, но только там, где не присутствуют годичные утолщения ксилемы. Кстати, там же наблюдается большее, чем обычно, число митохондрий. Удивительно, но число передаточных клеток в 25-миллиметровой зоне от кончиков корней кукурузы и фасоли заметно возрастает после действия умеренного засоления (возможно, для удаления большего количества Na+ из ксилемы). Если выведение Na+ из ксилемы представляет собой важный элемент адаптации к засолению, куда же этот Na+ девается? Некоторое количество может накапливаться в вакуолизированных клетках (например, в завершивших рост корнях в основании стебля и вдоль удлиненного стебля). Так, концентрация Na+ в ксилемном соке у ячменя, выращиваемого при 100 мМ NaCl, снижается от 10 мМ вблизи основания стебля и до 2 мМ у 8-го листа. Это снижение, равное примерно 1 мМ на одно междоузлие, соответствует удалению (выведению) примерно 3 мМ Na+ в сутки. К 45-му дню общая концентрация Na+ повысилась до 350 мМ/г сырой массы.
Таким образом, выведение Na+ из транспирационного тока и его накопление в очень высоких концентрациях в тканях междоузлий защищает молодые растущие ткани. Этому также содействует быстрый рост ячменя, благодаря чему новые ткани как бы «убегают» от надвигающегося «солевого фронта».
Рециркуляция натрия во флоэме и компартментация в побеге. Рециркуляция Na+ обратно в корни по флоэме до уровня, значительно влияющего на общее его накопление в листе, установлена на многих культурных растениях (люпин, клевер, кукуруза, сладкий перец), причем размер рециркуляции связан с солевыносливостью культуры. Например, он оказался выше у солевыносливого дикого вида томатов в сравнении с культурным. Размер и направление рециркуляции зависят от многих условий. Например, растения сладкого перца, выращиваемые по технологии расщепленной корневой системы, не транспортируют Na+ из побега в корень при засолении, но делают это при удалении NaCl из корнеобитаемой среды. Это же происходит при нанесении NaCl на лист («солевая» подкормка) в варианте без засоления, в результате чего Na+ обнаруживается в совершенно разных, более мобильных компартментах, чем было бы при выращивании в варианте с засолением.
Защита молодых листьев представляет собой ключевой элемент солевыносливости культур. Это обусловлено скорее всего малочисленностью вакуолей в этих типах клеток (для удержания Na+) и чувствительностью к Na+ ростовых процессов, особенно синтеза белка. Часто отмечалась низкая концентрация Na+ в молодых листьях, что объяснялось как низкой скоростью транспирации, так и коротким периодом жизни. Однако это не исключает возможности предпочтительного удаления Na+ из ксилемы и флоэмы, питающих эти ткани.
Очевидно, что Na+ избирательно удаляется из растворов, движущихся к органам, которые растение стремится защитить. Этот Na+ или запасается в месте удаления из ксилемы (о чем говорилось выше), или направляется в органы, которыми побег готов пожертвовать, например в более старые листья. Результат может быть достигнут путем движения вниз по флоэме (базипетально) и вверх по ксилеме (акропетально) с результирующим потоком Na+ (нетто-поток), направляющимся в строго определенную часть побега. Такой интенсивный круговорот растворов по ксилеме и флоэме описан для других растворов, в частности С- и N-содержащих веществ на горохе, а также для Na+, целенаправленно транслоцируемого в листья-мишени. Более того, показано избирательное движение растворов по флоэме, в частности 22Na+, экспортируясь из донорного листа, не поступает в молодые растущие ткани ни корня, ни побега. Это, вероятно, обусловлено избирательным выведением Na+ из флоэмы до достижения им молодых частей побегов.
Аналогично в пределах листа показано предпочтительное накопление Na+ в эпидермальных клетках, предписываемое повышенной активности неселективных катионных каналов в плазма-лемме этих клеток. Можно ожидать, что накопление Na+ в более вакуолизированных, метаболически менее активных клетках эпидермиса будет иметь адаптивное значение. Высокая концентрация Na+ в клетках обкладки свидетельствует о том, что этот тип клеток тоже может быть вовлечен в уменьшение накопления Na+ в ключевых фотосинтетических клетках мезофилла листа.