Рост растений в условиях засоления
Возможные причины лимитирования роста при засолении. Одним из центральных является вопрос о том, что более чувствительно к засолению — рост клеток растяжением или их деление, т. е. является ли причиной более низкой скорости роста листа меньший конечный размер клеток или меньшее их число в листе. В основе роста клеток лежит новообразование конституционных веществ, для которого в первую очередь необходимы азот, углеродсодержащие соединения, а также АТФ.
Поступление же азота может быть меньше при хлоридном засолении, так как Cl- ингибирует поглощение NO3-. Маловероятно, что низкая скорость поглощения азота может объяснить быстрый ответ растения на засоление, но полностью исключить этого нельзя. Рост действительно реагирует быстро на изменение концентрации NO3- в корнеобитаемой среде. Так, OCP растений салата снижалась в 2 раза в течение двух дней после удаления NO3- из питательного раствора. Скорость синтеза белка при хлоридном засолении, несомненно, ниже, чем без засоления. Однако прямых данных о том, что именно это вызывает ингибирование роста, нет. Уменьшение синтеза белка может быть скорее следствием торможения роста растяжением, нежели причиной.
При засолении, особенно хлоридном, наряду с торможением поступления NO3- происходит ингибирование восстановления нитратов, в результате значительно повышается содержание растворимых питательных веществ в корнях. Под действием Na2SO4 (в отличие от NaCl) повышается содержание общего, белкового и нитратного азота, глутамина, но снижается содержание небелкового, аминного азота и аспарагина. Возможно также накопление значительного количества NH3, что, по мнению некоторых исследователей, может вызвать аммиачное отравление и соответственно угнетение растений.
Концентрация углеводов (как растворимых, так и нерастворимых) в результате действия засоления, ингибирующего рост, возрастает или остается без изменения и в корнях, и в побеге. Поэтому этот фактор вряд ли может быть причиной ингибирования роста. ИФ при засолении обычно снижается. Поскольку концентрация солей в листьях при этом возрастает, снижение фотосинтеза можно было бы объяснить токсичностью солей. Однако данные о причинной связи концентрации ионов и ИФ отсутствуют. Напротив, после удаления NaCl фотосинтез восстанавливается намного быстрее, чем нормальный ионный статус листа. Причины ингибирования фотосинтеза могут быть иными, связанными, например, с ускорением старения листьев или низкой активностью акцептора ассимилятов из-за подавления роста.
Энергообеспеченность процессов роста не связана с поступлением углеводов, так как скорость синтеза АТФ может быть лимитирующей даже при их избытке.
Определим, действительно ли скорость синтеза АТФ при засолении должна быть намного выше, чем в отсутствие засоления. Представляется, что дополнительные затраты на транспорт NaCl и запасание в вакуоли при засолении достаточно значительны, но не чрезмерны и могут возрасти в корнях на 50—70% (по расчетным данным). Имеются также данные, что потребность в АТФ на регуляцию ионного статуса у галофитов, быстро растущих при засолении, составляет 10—20% от общего количества АТФ, синтезируемого в растении. Для негалофитов, растущих при гораздо меньших концентрациях солей, потребность в АТФ, вероятно, намного ниже. Интересно, что ИД сорго при засолении была на 6% ниже, чем в контроле. Однако ИД не может служить показателем скорости синтеза АТФ, так как у большинства неадаптированных к засолению видов весомый вклад в дыхание вносит цианидрезистентное дыхание, не сопряженное с синтезом АТФ.
На скорости синтеза АТФ теоретически могут сказаться также нарушения фосфорного обмена, причем характер нарушений зависит как от особенностей метаболизма растений, так и от типа засоления. В условиях хлоридного засоления у одних видов растений поглощение фосфатов из почвы подавляется незначительно, но резко ослабляется передвижение поглощенного фосфата из корней в надземные органы. У других видов подавление поглощения фосфора из почвы выражено значительно сильнее, в то время как снижение поступления его в растение выражено достаточно слабо. В условиях сульфатного засоления накопление фосфора в тканях растения уменьшается. Однако во всех случаях не весь фосфор используется растением и накапливается в неорганической форме. Отрицательное влияние засоления на фосфорный обмен сильнее выражено на ранних этапах онтогенеза в период активных ростовых и формообразовательных процессов. Таким образом, маловероятно, что доступность Pн в тканях при засолении может ингибировать синтез АТФ. Чтобы противостоять низкому водному потенциалу засоленного субстрата, некоторые растения используют поглощенные соли для осмотической адаптации. В клетках ионы солей компартментируются в вакуоли, а осмотический баланс между вакуолью и цитозолем поддерживается за счет накопления совместимых с цитоплазмой органических соединений (сахарозы, сорбита, маннита, пролина, бетаина и др.). В частности, пролин и бетаин способствуют стабилизации ферментов. Скорость синтеза бетаина и оборота белка при стрессе резко возрастают, причем концентрация бетаина достигает 100 мМ/г сухой массы, что соответствует содержанию его у галофитов. Тем не менее есть определенная метаболическая цена синтеза бетаина (2 мг гексозы/г в сутки) и оборота белка.
Об увеличении относительной доли дыхания от фотосинтеза свидетельствуют также данные, согласно которым при сильном засолении (150 моль NaCl/м3) доля ночного дыхания 4-го листа ячменя составила 14% от дневной ассимиляции, тогда как у контрольных растений (0 моль NaCl/м3) и при двукратном снижении засоления (75 моль NaCl/м3) была вдвое ниже.
Однако снижение роста нельзя полностью объяснить повышением только дыхательной цены жизни в условиях засоления. Важной реакцией растений на засоление является изменение распределения углерода — увеличение фондов неструктурных углеводов, недоступных для роста и дыхания, но участвующих в поддержании тургора. У адаптированных к произрастанию на засоленных почвах галофитов эту функцию выполняют соли.
Осмотическая адаптация, затраты энергии и рост. В связи со сравнительно-физиологическими аспектами осмотической адаптации при дефиците влаги и засолении возникает вопрос о метаболических затратах на подобную адаптацию, в том числе затратах на выведение солей, их внутриклеточную компартментацию и выделение через солевые железы. Оказалось, что они относительно невелики в сравнении с затратами, необходимыми для синтеза органических веществ для осмотической адаптации. Число молей АТФ, потребляемых при использовании 1 моля NaCl в качестве осмотика, составляет 4 в клетках корня и 7 в клетках листьев. Вместе с тем энергетические затраты на биосинтез и аккумулирование растворов в листьях почти на порядок выше: 34 моль АТФ для маннита, 41 — для пролина, 50 — для глицин-бетаина и около 52 — для сахарозы. Эти расчеты сделаны исходя из предположений, что на 1 фотон падающего света образуется 0,5 моль АТФ, а источником азота служат нитраты.
Однако до сих пор не ясно, образуют ли выращиваемые при засолении растения (в сравнении с незасоленным вариантом) больше или меньше органических активных веществ при одном и том же значении водного потенциала. Так, у ячменя, выращиваемого при действии ПЭГ и NaCl, накопление пролина и глицина при равном значении осмотического давления было примерно одинаковым. Однако эта тенденция относится только к азотсодержащим компонентам. Так, в аналогичных условиях выращивания у генотипов томатов содержание аминокислот в листьях было одинаковым, а содержание растворимых сахаров выше у растений, выращиваемых на ПЭГ.
Логично предположить, что если аккумулирование органических растворов требует больше энергии, чем накопление ионов, растения при одинаковых значениях водного потенциала должны расти быстрее на засоленной, чем на сухой почве. Однако опыты не дают однозначного ответа. Так, в вышеупомянутых опытах с ячменем растения росли лучше в ПЭГ, чем при засолении, в то время как генотипы томатов росли также или даже хуже в ПЭГ, чем при засолении. Сравнение вариантов с NaCl и со смесью солей, не содержащих NaCl, при одинаковом осмотическом давлении показало, что даже солеустойчивый ячмень рос лучше в среде без NaCl, как и неустойчивые кормовые бобы. Однако во всех этих опытах отсутствовали Ca2+ и соответственно индуцированный NaCl дефицит кальция. В присутствии NaCl активность Ca2+ снижается, а в присутствии ПЭГ нет, а в смеси солей (например, растворе Xoгланда) концентрация Ca2+ была достаточно высокой.
Несмотря на то что корни находятся в зоне непосредственного действия засоления и засухи, их рост ингибируется меньше, чем рост побегов. Ионный статус корней также более благоприятен и, что особенно важно, концентрация ионов в них не возрастает по мере усиления действия фактора засоления, как у листьев, и концентрация Na+ и Cl- в них зачастую ниже, чем у внешнего раствора, что редко наблюдается у листьев. Так, у пшеницы, выращиваемой при 150 мМ NaCl, концентрация Na+ в корнях колебалась в зависимости от генотипа в пределах 20—40 мМ на фоне низкой концентрации СИ и K+. Поскольку концентрация K+ в корнях, как правило, ниже, чем в побегах, возникает закономерный вопрос о роли органических растворов в осморегуляции. Однако отсутствуют данные об их значительном накоплении в корнях. Более того, их концентрация в корнях ниже, чем в листьях. Так, концентрация пролина и глицин-бетаина в расчете на сухую массу в корнях ячменя в диапазоне засоления 100—200 мМ NaCl в 5 раз ниже, чем в побегах на фоне опять-таки относительно более низких концентраций Na+ и Cl-. Аналогичная тенденция наблюдается также при выращивании в ПЭГ.
Интересно, что рост корней кукурузы в длину при действии засоления 100 мМ NaCl полностью восстанавливался через 24 ч, а при 150 мМ NaCl — через 36 ч. Вместе с тем постепенное повышение концентраций NaCl вплоть до 100 мМ не оказывало влияние на рост корней, а солевой шок сразу же ингибировал рост, что, видимо, обусловлено плазмолизом клеток корня. Через двое суток после солевого шока при 250 мМ NaCl концентрация Na+ и Cl- в побеге составила 3,5 мМ/г, а при постепенном повышении засоления — лишь 1,4 мМ/г сухой массы. Интересно, что благодаря выходу из корней концентрация калия там уменьшилась в 4 раза.
Кинетика ростовых процессов при засолении. Растения реагируют на засоление относительно быстро (минуты-часы) и медленно (сутки и даже месяцы). Рассмотрим специфику быстрой и медленной реакций растений в разных временных диапазонах.
Минуты-часы. Листья отвечают на быстрое изменение режима засоления в лабораторных условиях таким же мгновенным изменением скорости роста (рис. 10.6). Быстрое уменьшение скорости растяжения листа в ответ на резкое увеличение засоления в корнеобитаемой среде отмечено для кукурузы, риса, пшеницы, ячменя и других культур. Подобный характер реакции наблюдается и после добавления KCl или ПЭГ, что свидетельствует о соленеспецифичности реакции. Как быстрое падение скорости роста после начала действия высокой засоленности, так и мгновенное ее восстановление после снятия действия стрессора указывают на вовлеченность в эти процессы исключительно водного статуса клеток. Уже через несколько минут после первоначального снижения скорости роста клеток начинается постепенное ее восстановление, длящееся 30 мин или более, до достижения нового стационарного состояния. Время восстановления и скорость роста в новом стационарном состоянии зависят от уровня засоления и условий окружающей среды.
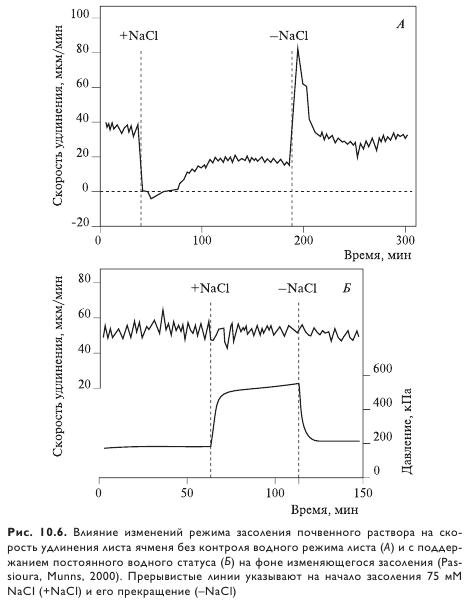
Ростовая функция корней реагирует на воздействие солевого стресса качественно так же, за исключением более быстрого восстановления после начала действия стрессора. При умеренном осмотическом стрессе (0,1—0,4 МПа маннитола или KCl) скорость удлинения корней кукурузы практически полностью восстанавливается в течение 1 ч, а при более сильном засолении NaCl (0,7 МПа) в течение 24 ч. Интересно, что характер ростовой реакции корней зависит от того, присутствует ли в корнеобитаемой среде Ca2+ в момент действия засоления. Например, удлинение корней у кукурузы не тормозилось при засолении 80 мМ NaCl в присутствии Ca2+ в концентрации 10 мМ. В отсутствии же Ca2+ рост тормозится и последующее его восстановление происходит достаточно медленно. Удлинение корней сорго при концентрации NaCl 150 мМ тормозилось на 20% в присутствии Ca2+ и на 80% в его отсутствие.
Сутки. К этому времени скорость роста листьев и корней, как правило, стабилизируется на более низком стационарном уровне, причем рост листьев тормозится сильнее, что особенно заметно на сухих почвах. Концентрация Na+ и Cl- в растущих листьях в этот момент гораздо ниже токсичного уровня. Так, у пшеницы, выращиваемой при 120 мМ NaCl, при 25%-м ингибировании скорости роста листьев, максимальная концентрация Na+ в растущих листьях составила 20 мМ, а Cl- — 60 мМ (рис. 10.7). Аналогичная тенденция отмечается и для корней.
Таким образом, возможно именно водный стресс является ключевым при торможении роста. Однако есть целый ряд данных, указывающих на то, что в суточном интервале времени скорее гормональные сигналы, а не водный стресс, контролируют рост на засоленных почвах. В частности, растяжение листа на засоленной почве в течение 24 ч не реагировало на улучшение водного статуса растения.

В последнем опыте растения выращивали в сосудах, которые затем помещали в камеру давления, где растения поливали соленой водой. Затем в камере создавали давление, равное осмотическому давлению раствора NaCl с концентрацией 100 мМ. При этом не было обнаружено никакого эффекта указанного давления на рост контрастных по солеустойчивости видов (ячмень, пшеница, люпин, египетский клевер, галофиты) в течение 8 сут. Эти данные подтверждают высказанное предположение о роли гормонального фактора в регуляции роста при засолении.
У очень солечувствительных видов солеспецифические эффекты можно заметить через несколько суток после действия сильного засоления (например, у люпина при концентрации NaCl свыше 100 мМ). Токсичность солей хорошо заметна прежде всего на более старых листьях, транспирирующих гораздо больше, чем молодые листья. Визуально симптомы токсичности можно заметить через несколько суток или даже недель, что зависит от уровня засоления, окружающих условий (температура, относительная влажность, состав питательной смеси) и генотипической способности корней воспрепятствовать проникновению солей в транспирационные токи.
Недели. Результаты действия засоления на растения могут стать явными через несколько недель, особенно у более чувствительных видов, характеризующихся высокой скоростью поглощения солей и неэффективной компартментацией их в листьях. Повреждения проявляются в виде пожелтения или даже гибели относительно более старых листьев. У солеустойчивых видов, у которых ниже скорость поглощения солей и выше способность компартментации солей в вакуолях клеток, предотвращается их накопление в цитоплазме или в клеточных стенках. В результате процессы развития и метаболическая активность у этих видов примерно такие же, как и при водном стрессе.
Месяцы. У однолетних культур засоление влияет на формирование и жизнеспособность репродуктивных органов. У зерновых оно снижает число цветков в колосе, изменяет время цветения и созревания. Подобное явление имеет место и при водном стрессе. Однако концентрации Na+ и Cl- в репродуктивных органах (как было показано на сортах пшеницы и ячменя разной солеустойчивости) достаточно низкая, чтобы оказать негативное влияние на метаболизм. Кроме того, большинство клеток в репродуктивном апексе невакуолизированы и любая поступающая в них соль аккумулируется в цитоплазме. Поэтому транспорт Na+ и Cl- по флоэме в апекс должен находиться под строгим контролем, чтобы предотвратить их концентрирование до токсичного уровня.
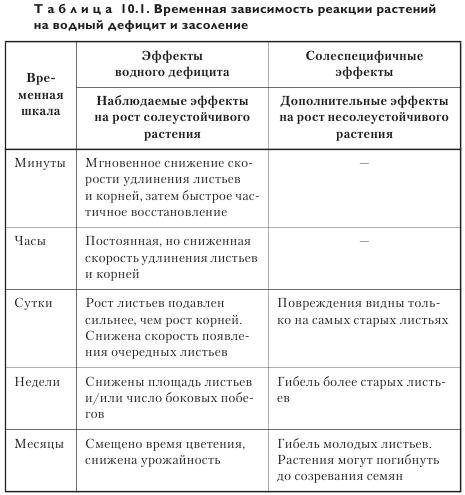
В таблице 10.1 показана последовательность событий, происходящих в растении после действия засоления.
Таким образом, временная динамика реакции на засоление выглядит следующим образом:
• через несколько минут клетки теряют воду и сморщиваются;
• через несколько часов клетки вновь восстанавливают свою первоначальную форму, но скорость растяжения клеток уже снижена, что приводит к уменьшению скорости роста листьев и корней;
• по прошествии нескольких суток изменения в делении и растяжении клеток ведут к снижению скорости появления очередных листьев и их конечных размеров. При этом рост листьев ингибируется сильнее, чем рост корней. У растений с высокой скоростью поглощения солей наиболее старые листья демонстрируют симптомы повреждения;
• через несколько недель становится заметным ингибирование роста боковых побегов, и у растений с высокой скоростью поглощения гибнет целый ряд листьев. Вместе с тем скорость образования новых листьев у генотипов может и не различаться;
• по прошествии месяцев различия между растениями с разной скоростью поглощения солей становятся очевидными.