Влияние засоления на дыхание и углеродный баланс растений
При изучении концентрационной зависимости часто (особенно у культурных растений) наблюдали двухфазное изменение дыхания: при умеренном засолении дыхательная активность повышалась, при сильном — снижалась (по отношению к контролю). Это обусловлено главным образом соответствующим изменением дыхания для поддержания на фоне примерного постоянства эффективности превращения ассимилятов в биомассу. Следует, однако, отметить, что реакция дыхания зависела не только от уровня засоления, но и от солеустойчивости вида. Так, ИД молодых растений клевера (Medicago sativa L.) достигала максимума при увеличении концентрации NaCl до -0,3 МПа (в эквивалентах осмотического давления) и снижалась при больших концентрациях соли. Концентрация NaCl, эквивалентная -0,3 МПа, не влияла на скорость роста растений. У более устойчивых к засолению растений сахарной свеклы (Beta vulgaris L.) дыхательная способность листьев слабо реагировала на концентрации NaCl, при которых рост и фотосинтез подавлялись.
Дыхание листьев молодых растений ячменя и пшеницы при концентрации NaCl в питательном растворе 75 и 150 мМ в ночные часы было ниже у опытных растений. При этом концентрация хлорида в листьях ячменя и пшеницы составляла 170 и 120 мМ/г сырой массы при 75 мМ NaCl в питательном растворе. При 150 мМ NaCl концентрация NaCl в листьях ячменя достигала 420 мМ/г, а у пшеницы в фазе 5 листьев отмирали все листья. Даже при кратковременном изменении содержания соли в питательном растворе реакция дыхания отмечалась уже через 1 ч.
Компоненты дыхания (дыхание на рост и дыхание на поддержание) изменяются по-разному в зависимости от уровня солеустойчивости полевых культур и режима засоления (фасоль, кукуруза, люцерна). Дыхание поддержания с усилением засоления возрастало вплоть до концентрации, при которой стали визуально заметны повреждения листьев. Эта концентрация неодинакова для различных видов. При дальнейшем повышении засоления дыхание поддержания снижалось.
Воздействие засоления снижало как площадь листьев, так и ИФ единицы площади листа при одновременном увеличении ИД главным образом за счет дыхания поддержания листьев. Дыхательные затраты в расчете на целое растение снижались, что обусловлено падением дыхания роста.
Стационарное засоление оказывает следующее влияние на углеродный баланс целого растения:
• снижается фотоассимиляция CO2 в расчете на одно растение (единицу площади посева) из-за уменьшения площади листьев и ИФ;
• дыхание роста снижается пропорционально уменьшению фотосинтеза целого растения, параллельно со снижением уровня метаболической активности;
• метаболические затраты по компартментации и выведению солей возрастают, хотя их доля от общих дыхательных затрат невелика.

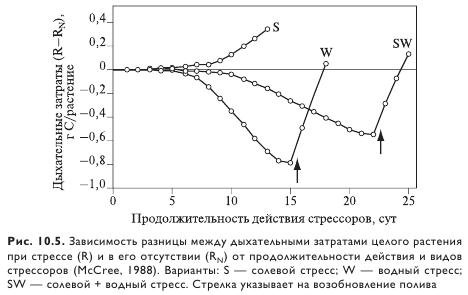
При умеренном, но хроническом солевом стрессе (осмотический потенциал -0,56 МПа) рост листьев растяжением у сорго тормозился, хотя растения были способны поддерживать тургор. При этом суточный прирост биомассы был меньше, чем в контроле без засоления, что указывает на заметные траты на адаптацию к засолению. Сочетание водного и солевого стрессов в одном опыте позволяет ориентировочно оценить эти затраты. При прекращении полива водный стресс у растений в варианте с засолением развивается медленнее, чем в варианте только с водным стрессом (рис. 10.3). В первом случае рост листьев был уже ингибирован и интенсивность транспирации была ниже. В результате растения оказались способны перенести более низкий водный потенциал, чем при только водном стрессе. Однако в обоих этих вариантах прирост биомассы на единицу ассимилированного CO2 была выше, чем в контроле с орошением (рис. 10.4). Следовательно, влияние фотоассимилятов на осмотическую адаптацию было меньше, чем на синтез биомассы (сравните варианты SW и S, а также W и N).

При анализе углеродного баланса весьма информативным может быть сравнение суточных дыхательных затрат растения при солевом стрессе и без него. Если затраты на адаптацию меньше, чем на синтез биомассы, растения при стрессе должны терять на дыхание меньше ассимилятов, чем растения в контроле. Оказалось, что у растений при водном стрессе дыхательные траты меньше, чем при сочетании водного и солевого стрессов. Более того, в обоих вариантах после полива величина суточных дыхательных затрат достигает исходного уровня (до наложения стресса). Это указывает на то, что все запасенные фотоассимиляты быстро используются после начала полива.
Растения при засолении характеризуются наибольшими суммарными дыхательными затратами, что свидетельствует о дополнительных тратах на адаптацию в случае действия только солей. Действительно, дыхательные потери растения при стрессе на 2% больше, чем в норме и имеют тенденцию к увеличению по мере возрастания продолжительности действия засоления.
При сочетании водного и солевого стрессов, напротив, суммарные дыхательные затраты растения меньше, чем у контрольных растений (R - RN < 0), т. е. растения при наложении на солевой стресс водного воспринимают ситуацию уже в реализованном состоянии адаптации (рис. 10.5).