



Минеральное питание растений при дефиците воды
При неограниченном доступе элементов питания и, соответственно, отсутствии лимитирования накопления биомассы ЭИВ достигает наибольших величин. Это относится ко всем основным макроэлементам и особенно к азоту, причем ЭИВ зависит от формы азотного питания (табл. 6.3).
ЭИВ меньше при использовании симбиотического, чем аммонийного и нитратного азота. Среди двух последних форм ЭИВ ниже на аммонийном, чем на нитратном фоне у двудольных культур, и примерно одинакова у злаков. Отмеченные меньшие величины ЭИВ на симбиотическом и аммонийном азоте были обусловлены главным образом низкой относительной скоростью накопления биомассы, что указывает на тесную корреляцию этих параметров. Механизмы, с помощью которых разные формы азотного питания могут влиять на устьичную проводимость, включают активное поступление нитратов в замыкающие клетки. Однако в условиях нормального азотного питания доминирующим неорганическим анионом, участвующим в открывании устьиц, является хлорид (а не нитрат). Соответственно, нитратный переносчик может и не обусловливать различия в функционировании устьиц растений в связи с нитратной и аммонийной формами азотного питания. В связи с этим представляют интерес данные о том, что gs листьев растений пшеницы, выращиваемой на аммонийном азоте, была на 20—40% ниже, чем на нитратном азоте, причем различия достигали наибольших значений при высокой засоленности почв.
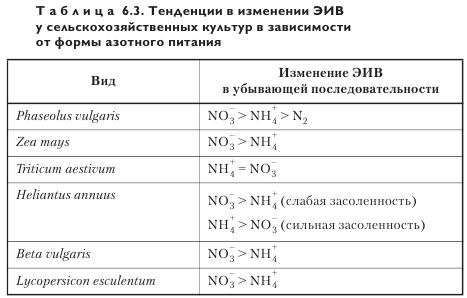
Теоретические расчеты потерь воды на накопление органического азота в биомассе (моль Н2O/моль ассимилированного N) в зависимости от источника азотного питания (NH4+, NO3 или N2) впервые были проведены более 20 лет назад. Полученные данные оказались выше в варианте с N2 и NO3 независимо от сделанного допущения о месте восстановления NO3 — в корнях или побеге. Что касается экспериментальных данных, полученных на подсолнечнике и фасоли, потери воды на ассимиляцию углерода заметно отличаются от теоретических расчетов. Так, они выше (ЭИВ ниже) для фасоли при выращивании на N2 (по сравнению с NH4+). Модель предполагает, по меньшей мере, одинаковые величины потерь воды на ассимиляцию углерода, в то время как в опытах они оказываются ниже в варианте с NO3 (по сравнению с NH4+). Объяснение этого факта сводится или к более низкому АТО, или к большим дыхательным потерям органического углерода. Возможно и сочетание обоих факторов. Важно подчеркнуть, что относительная скорость роста (OCP) как у фасоли, так и у подсолнечника ниже в варианте с аммонийным азотным питанием, чем нитратным, а у пшеницы OCP практически одинакова. Таким образом, относительно меньшая, равная или большая OCP в варианте с NH4+, чем с NO3, не определяет величину транспирационных потерь воды на ассимиляцию углерода. Более того, ATO у медленно растущей пшеницы на низком фоне NO3 меньше, чем на фоне оптимального обеспечения NO3 (данные по другим формам азотного питания, к сожалению, отсутствуют). Чтобы понять, каким образом зависимость OCP от форм азотного питания влияет на АТО, необходимы дополнительные исследования. Поэтому пока преждевременно искать механизмы для объяснения более высокого соотношения устьичной и мезофильной проводимости в варианте с NH4+ (по сравнению с NO3).
Следует заметить, что механистические связи между нетто-поглощением питательных веществ и воды достаточно сложны, а линейная зависимость между этими параметрами скорее исключение, чем правило. Именно к подобным исключениям относится наблюдаемая корреляция между содержанием кремния в растениях ячменя и пшеницы и количеством транспирируемой воды. По сути это предполагает поглощение воды и кремниевой кислоты в тех же пропорциях, в каких они находятся в почвенном растворе и указывает на возможность использования характера аккумуляции кремния в качестве индикатора ЭИВ.
Следует также иметь в виду, что минеральное питание может влиять на ЭИВ через изменение плотности устьиц, размеров их апертуры и, соответственно, устьичной проводимости. В частности, снижение скорости транспирации при дефиците калия было полностью обусловлено меньшей плотностью устьиц.
Другой катион — Ca2+, благодаря концентрированию в апопласте эпидермиса листа, участвует в регуляции открывания устьиц. Влияние кальциевого питания на устьичную проводимость и ЭИВ изучалось как на кальциефилах (люпин), так и на кальциефобах. Высокая концентрация Ca2+ в ризосфере у последних снижала сопряженность gs с ИФ и соответственно ATO благодаря повышению концентрации Ca2+ в апопласте листа. Что касается кальциефилов, данные о снижении концентрации Ca2+ в клетках мезофилла и трихомах листьев свидетельствуют о том, что эпидермальный апопласт может иметь «нормальную» концентрацию Ca2+ и, соответственно, демонстрировать «нормальную» функцию устьиц, несмотря на высокое содержание Ca2+ на уровне целого растения. В других опытах, где дефицит кальция повышает ПТ, путем кратковременного изменения текущих значений фотосинтеза и транспирации листьев показано, что фотосинтез подавляется сильнее, чем транспирация. В таких случаях любое влияние дефицита питания на потерю углерода при выделении, секреции, дыхании и потере органа объясняет изменение ЭИВ на количественном уровне.