



Механизмы детоксикации АФК
На сегодняшний день известны следующие методы определения уровня и локализации окислительного стресса в растениях:
• определение активности антиоксидантных ферментов;
• определение концентрации АФК: супероксид-ионов, перекиси водорода и др.;
• измерение концентрации маркеров окислительного стресса: малонового альдегида, окисленного гуанозина и др.;
• гистохимические методы определения локализации АФК;
• прямое измерение концентрации АФК методом парамагнитного резонанса;
• масс-спектрометрические методы измерения количества АФК в клетке.
Определение и классификация антиоксидантов. Антиоксиданты (АО) — вещества, снижающие активность радикальных окислительных процессов по следующим механизмам:
• ингибирование радикальных форм активных кислородных метаболитов, способных инициировать образование органических радикалов;
• изменение структурной организации субстрата, замедляющее окисление;
• прерывание окислительной цепи посредством взаимодействия с органическими радикалами;
• снижение концентрации кислорода;
• связывание или окисление ионов металлов переменной валентности, инициирующих разложение перекисей и образование радикалов;
• перевод перекисей в стабильные продукты окисления: спирты, альдегиды, кетоны и др.
Выделяют также антиоксиданты вторичной защиты, которые обеспечивают восстановление от повреждений после действия АФК. К ним относят процессы репарации ДНК, гидролитические ферменты, осуществляющие «выщепление» окисленных липидов из мембраны, протеолиз поврежденных белков, комплекс реакций, связанных с метаболизацией продуктов ПОЛ. Детоксикация в широком смысле также включает замещение поврежденных молекул ДНК, белков, мембранных липидов комплексной системой интегрированных репарирующих ферментов и процессов замещения. Однако непрерывная работа процессов репарации приведет к их инактивации, так как они функционируют также в качестве мишеней АФК.
Удобно характеризовать группы антиоксидантов в зависимости от их молекулярных масс, причем первую группу составляют низкомолекулярные соединения, а вторую — высокомолекулярные ферменты, белки и пептиды, способные связывать ионы металлов переменной валентности.
Низкомолекулярные антиоксиданты. К первой группе относятся разнообразные соединения: глутатион, аскорбат, токоферолы, каротиноиды, полиамины, некоторые аминокислоты и др. Отличительной особенностью большинства эндогенных низкомолекулярных антиоксидантов является нелинейная зависимость между их концентрацией и степенью ингибирования свободнорадикальных процессов.
Антиоксиданты можно подразделить на две группы: водорастворимые протектанты и антиоксиданты липидной фазы.
Из водорастворимых антиоксидантов наиболее эффективными являются глутатион и аскорбиновая кислота, обнаруженные практически во всех участках клетки, но в особенно больших концентрациях — в хлоропластах и апопласте. Защитное действие глутатиона связано с окислением его SH-группы, приводящим к димеризации в дисульфид. Антиоксидантные свойства аскорбата основаны на функционировании одноэлектронных циклических переходов между гидро-, семидегидро- и дегидроаскорбатными формами, чему способствуют подвижность протонов. Скорость этих превращений зависит от металлов переменной валентности, других пар окислитель-восстановитель и pH среды.
Антиоксидантный эффект аскорбиновой кислоты и глутатиона главным образом реализуется посредством участия в работе ферментативных антиоксидантов, особенно ферментов аскорбат-глутатионового цикла. Кроме того, они способны непосредственно ингибировать АФК, легко перехватывая O2', HO2', RO2', HO', 1O2 и др.
Аскорбиновая кислота (витамин С) и восстановленный глутатион (GSH), являясь гидрофильными антиоксидантами, находятся в водной фазе клетки, в хлоропластах, митохондриях и других структурах, а также в межмембранном пространстве клеточных органелл. Глутатион, являясь полифункциональным соединением, представляет собой основной восстановитель клетки. Как и аскорбиновая кислота, он восстанавливает токофероксильные радикалы, а также участвует в обезвреживании вторичных метаболитов окислительного обмена.
Важной функцией водорастворимых соединений является их роль в восстановлении других низкомолекулярных АО. Так, клеточный GSH участвует в поддержании пула восстановленного аскорбата. Аскорбиновая кислота способна восстанавливать мембранно-связанный токоферол, а также служит высокоспецифичным донором в реакции эпоксидации виалоксантина.
Кроме непосредственно антиоксидантного действия, глутатион и аскорбат выполняют многочисленные функции в регуляции метаболизма растений, в частности, участвуют в поддержании окислительно-восстановительного потенциала.
Антиоксиданты липидной фазы представлены двумя основными классами низкомолекулярных соединений. К первому относят фенольные токоферолы и близкие к ним по строению убихиноны и витамин К. Гидрофобные антиоксиданты предохраняют от разрушения мембраны. Так, локализующийся в мембранах а-токоферол (витамин Е) вызывает обрыв цепей свободнорадикального окисления путем взаимодействия с пероксильными и алкоксильными радикалами. Таким способом токоферол нейтрализует свободные радикалы мембранных липидов, перехватывая и устраняя их. Углеводородная цепь токоферола, встраиваясь в мембрану, образует ассоциации с ненасыщенными жирными кислотами мембранных липидов. Токоферол взаимодействует с O2' и 1O2. Кроме того, токоферолы снижают проницаемость мембран, а также связывают свободные жирные кислоты, избыток которых дестабилизирует мембранную структуру.
Помимо токоферола высокую антиокислительную активность проявляют другие фенольные соединения, в частности полифенолы, служащие донорами водорода. Многие из них способны связывать металлы переменной валентности, которые служат катализаторами свободнорадикального окисления. Из полифенолов как антиоксиданты наиболее известны различные флавоноиды.
К липофильным антиокислителям относятся каротиноиды, которые наиболее эффективны в тушении избыточной энергии триплетных хлорофиллов и синглетного кислорода. Воспринимая энергию возбуждения, они рассеивают ее в виде тепла, предотвращая тем самым возможность образования синглетного кислорода. Кроме того, каротиноиды могут взаимодействовать с органическими радикалами жирных кислот, действуя при этом в качестве не доноров водорода, а «ловушек» радикалов. В итоге цепь окисления обрывается. Антиокислительное действие каротиноидов видимо дополняет действие токоферолов, поскольку каротиноиды эффективны в субклеточных фракциях с низким уровнем кислорода, а токоферолы — с более высоким. Возможны и прямые взаимодействия между каротиноидами и токоферолами.
Фенольные соединения в основном перехватывают перекисные и алкоксильные радикалы, но могут ингибировать O2', HO2', 1O2, ОН'. Взаимодействие фенольных АО с органическими радикалами приводит к образованию феноксильных радикалов, которые в дальнейшем могут образовать хинолидные перекиси. Распад последних приводит к образованию хинонных форм молекул, которые не обладают антиоксидантными свойствами и восстанавливаются с помощью других АО, в частности аскорбата. Фенольные АО также участвуют в ингибировании или инактивации ферментов, активирующих кислород. Механизмы этих процессов могут быть основаны на конкуренции фенолов с субстратом при связывании с регуляторным центром фермента или инактивации активного центра.
Второй класс липорастворимых АО составляют ретинол, его предшественники и производные. Антиоксидантная роль каротиноидов проявляется в основном в ингибировании синглетного кислорода, но они могут обезвреживать и пероксирадикалы. Наилучшим эффектом обладают (3-каротин и ликопин, способные хелатировать ионы Fe2+, а также тушить триплетное состояние хлорофилла, являющегося одним из источников образования 1O2.
Скорость ПОЛ существенно зависит от структурной организации фосфолипидов мембран: чем плотнее их упаковка, тем меньше скорость окисления. Способностью стабилизировать липиды мембран обладают токоферолы, витамин К, убихиноны, каротиноиды, холестерол, другие соединения. Встраиваясь боковыми цепями между НЖК мембран, эти вещества образуют комплексы с их двойными связями.
Обнаружены AO-свойства низкомолекулярных стероидов (витамина D3 и экдистерона), моносахаридов, некоторых дипептидов (карнозина, карцинина, L-глутаминилгистамина и др.). Они могут взаимодействовать с активными радикалами, восстанавливать гидроперекиси, хелатировать металлы переменной валентности.
Системы антиоксидантной защиты. Растения обладают достаточной устойчивостью к окислительным повреждениям, которые возникают лишь при резком изменении физиологического состояния их организма. Это обусловлено существованием в растительной клетке эффективных антиоксидантных систем (АОС), способных обеспечить защиту как от кислородных радикалов, так и от синглетного кислорода.
Как восстанавливающий агент, аскорбиновая кислота способна напрямую реагировать с супероксид- и гидроксильным радикалами, однако основная ее функция в процессе ПОЛ состоит в регенерации токофероксильного радикала:

где AKH — аскорбиновая кислота; А — радикал монодегидроаскорбиновой кислоты.
Дальнейшее окисление монодегидроаскорбата приводит к образованию дегидроаскорбата. Окисленные формы аскорбиновой кислоты восстанавливаются в ферментативных реакциях монодегидроаскорбат- и дегидроаскорбатредуктазами.
Глутатион представляет собой основной восстановитель в клетке. Окисление сульфогруппы глутатиона (Г-SH) приводит к образованию окисленного глутатиона (Г-SS-Г), в котором две молекулы T-SH связываются, образуя дисульфидный мостик. В ходе окислительного стресса количество Г-SS-Г резко увеличивается, затем активируется синтез восстановленных форм глутатиона. Глутатион — полифункциональное соединение. Как и аскорбиновая кислота, он восстанавливает токофероксильные радикалы:

Он также участвует в восстановлении H2O2, ROOH, обезвреживании вторичных метаболитов окислительного обмена. Глутатионзависимые ферменты работают во всех частях клетки. Антиоксидантный эффект селена также опосредован глутатионпероксидазами.
Гидрофобные антиоксиданты предохраняют от разрушения мембраны. Локализующийся в мембранах а-токоферол (витамин Е) вызывает обрыв цепей свободнорадикального окисления путем взаимодействия с пероксильными и алкоксильными радикалами:

где ТОН — токоферол; ТО — токофероксильный радикал.
Образовавшийся токофероксильный радикал восстанавливается до токоферола или вступает в реакцию с новыми пероксильными радикалами: ТО' + ROO' —> молекулярные продукты. При этом образуется большое число молекулярных продуктов с эпокси-, кето-, гидроперокси- и другими группами. Таким образом, токоферол нейтрализует свободные радикалы мембранных липидов, перехватывая и устраняя их. Углеводородная цепь токоферола, встраиваясь в мембрану, образует ассоциации с ненасыщенными жирными кислотами мембранных липидов. Токоферол взаимодействует с O2' и 1O2. Кроме того, токоферолы снижают проницаемость мембран, а также связывают свободные жирные кислоты, избыток которых дестабилизирует мембранную структуру.
Помимо токоферола высокую антиокислительную активность проявляют другие фенольные соединения, в частности полифенолы (как доноры водорода). Многие из них способны связывать металлы переменной валентности, которые служат катализаторами свободнорадикального окисления. Из полифенолов в качестве антиоксидантов наиболее известны различные флавоноиды.
Основную роль в снижении уровня АФК играют ферментные АОС: СОД, каталаза, пероксидаза, глутатионтрансфераза, фосфо-липидгидропероксидаза и др. СОД катализирует реакцию восстановления O2':

препятствуя накоплению супероксида кислорода. В активных центрах СОД растений содержатся ионы меди, цинка, железа или марганца. Так, Cu-Zn-СОД локализована в основном в цитоплазме, хлоропластах и пероксисомах эукариот, Мn-СОД — в митохондриальных мембранах, а Fe-СОД — в хлоропластах большинства растений. Механизм взаимодействия СОД с супероксидным радикалом точно не выяснен. Предполагается, что сначала одна молекула супероксида взаимодействует с активным центром фермента. При этом металл, входящий в активный центр, восстанавливается, а супероксидный радикал окисляется до молекулярного кислорода:

Затем при участии второй молекулы супероксидного радикала происходит обратное окисление металла. При этом супероксид восстанавливается до пероксида водорода:

В реакции детоксикации пероксида водорода

важную роль играют Fe-содержащие ферменты — каталаза и различные пероксидазы. Активность каталазы связана в основном с митохондриями, цитоплазмой и пероксисомами. В отличие от пероксидаз, которые функционируют только при относительно низком уровне пероксида, каталаза способна эффективно работать при высокой его концентрации.
Пероксидазы — обширная группа ферментов, катализирующих реакции окисления органического и неорганического субстрата с использованием пероксида водорода или органических пероксидов в качестве акцепторов электронов:

где XH — восстановленный субстрат; X — окисленный субстрат.
Функции пероксидаз разнообразны. Они участвуют в синтезе лигнина, катаболизме ауксина, связаны с процессами роста и растяжения и т. д. К важным функциям пероксидаз относится участие их в защите растения-хозяина от патогена, катализ окисления фенолов до хинонов, а также образование супероксид-аниона и пероксида водорода (оксидазная функция). Антиоксидантную защиту, связанную с восстановлением пероксида водорода, осуществляют главным образом глутатион- и аскорбатпероксидазы. В вакуолях растений, а также у организмов с низкой активностью аскорбатпероксидазы детоксикация пероксидов эффективно обеспечивается гваяколпероксидазой.
Глутатионпероксидаза, локализованная в цитоплазме, плазма-лемме и в матриксе митохондрий, разрушает пероксид водорода и органические гидропероксиды свободных жирных кислот, нуклеотидов, нуклеиновых кислот, белков:

Среди глутатионпероксидаз выделяют связанную с мембранами селен-зависимую форму, катализирующую в основном редукцию гидропероксидов фосфолипидов. Эта фосфолипидгидроксипероксид-глутатионпероксидаза восстанавливает ROOH жирных кислот в составе фосфолипидов. В строме хлоропластов образуется много пероксида водорода, и в его элиминировании участвует несколько сопряженных ферментативных систем.
Пероксид водорода разрушается аскорбатпероксидазой, две формы которой локализованы в хлоропластах, а две другие — в мембранах глиоксисом и в цитоплазме. Окисленный аскорбат восстанавливается аскорбатредуктазой, в реакции также участвует восстановленный глутатион. В свою очередь, глутатион регенерируется глутатионредуктазой, в результате из Г—SS—Г путем НАДФН-зависимого восстановления образуется Г—SH:
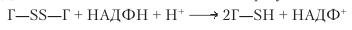
Эта реакция уменьшает или даже предупреждает прогрессирование ПОЛ и окислительный распад нуклеиновых кислот и белков. Глутатионтрансфераза восстанавливает только ROOH, причем один из ее изоферментов находится в хроматине и восстанавливает ROOH ДНК в ядре.
Гваяколпероксидазы, или «классические пероксидазы», катализируют окисление большого набора ароматических соединений с использованием пероксида водорода или органических пероксидов в качестве акцепторов электронов. Помимо участия гваяколпероксидаз в лигнификации, распаде ауксина и других процессах, им отводят важную роль и в защите клетки от окислительного стресса. Например, в вакуолях или клеточной стенке, не имеющих аскорбатпероксидаз, но содержащих в достаточно высоких концентрациях фенолы и аскорбиновую кислоту, может функционировать комплекс гваяколпероксидаза-фенолы-аскорбиновая кислота. При этом фенолы окисляются гваяколпероксидазой, а затем восстанавливаются аскорбиновой кислотой, в результате происходит детоксикация продуктов окисления фенолов. Таким образом, в клеточных стенках и вакуолях происходит эффективное обезвреживание пероксида водорода, что предотвращает возможность проникновения его в другие субклеточные структуры.
Рассмотрим последовательность реакций пероксидации и совместное действие ряда антиоксидантных систем в детоксикации продуктов ПОЛ.
На первом этапе пероксидации (А) окисляющий радикал X образует пентодиеновый радикал ненасыщенной жирной кислоты. Затем (B) образуются пероксильный радикал и конъюгированные диены — промежуточные продукты ПОЛ. На следующем этапе (В) пероксильный радикал перемещается к гидрофильной поверхности мембраны и, локализуясь на границе раздела фаз, взаимодействует с токоферолом (ТФ). В результате образуется токофероксильный радикал. На следующем этапе (Г) пероксильный радикал превращается в гидропероксид липида, ТФ восстанавливается аскорбиновой кислотой (АКН-). Далее (Д) ТФ возвращается в исходное состояние, а радикал аскорбиновой кислоты (AK ) регенерируется с помощью комплекса антиоксидантных ферментов. Репарация гидропероксида ненасыщенной жирной кислоты производится с помощью фосфолипазы A2 (Фл A2), которая вырезает поврежденную жирную кислоту из молекулы фосфолипида. Ее последующая детоксикация осуществляется при участии гваяколпероксидазы (ГП). Фосфолипидгидрокси-пероксидглутатионпероксидаза (Фл-Гп-ГП) производит детоксикацию гидропероксида ненасыщенной жирной кислоты в составе молекулы фосфолипида. Поврежденная жирная кислота замещается новой, которая поставляется в связанной с коэнзимом А форме (ЖК—KoA).
Вторичные метаболиты окислительных модификаций также обезвреживаются различными ферментами: глутатионтрансфераза катализирует конъюгацию с Г—SH ряда окисленных веществ, формальдегиддегидрогеназа и глиоксилаза, использующие Г—SH в качестве кофермента, окисляют свои субстраты до органических кислот, альдегиддегидрогеназа окисляет МДА, хинонредуктаза восстанавливает хиноны до дегидрохинонов, что предупреждает образование продуктов одноэлектронного восстановления семихинонов. Таким образом, ферментативная AOC обеспечивает разрушение не только АФК, но и их производных окисленных соединений.
При дефиците низкомолекулярных антиоксидантов (глутатиона, витаминов E и С) накапливаются АФК и нарастает содержание окисленных веществ, т. е. окислительный стресс возникает не только при избытке АФК и оксидативных модификаций макромолекул, но и при недостатке AOC (не только абсолютном, но и относительном). Следовательно, окислительный стресс — это сдвиг к преобладанию прооксидантов над антиоксидантами. Соотношение прооксидантов и антиоксидантов лежит в основе внутриклеточной редокс-регуляции. Эта концепция подразумевает признание регуляторных функций АФК и АОС. Итак, в регуляции биологических процессов следует выделить новую форму контроля — редокс-регуляцию, или контроль соотношения прооксидантов и антиоксидантов, по сути, определяющую возможность прогрессирования или блокады оксидативного стресса.
Образование АФК и последующие оксидативные изменения органических веществ представляют собой неспецифические реакции. У растений, устойчивых к различным воздействиям, процессы генерации АФК обычно заторможены, что во многом определяется активным функционированием у них систем детоксикации. Возможно, именно с этим связана в значительной мере и большая стойкость мембран устойчивых растений к стрессорам. Накоплен обширный экспериментальный материал, который свидетельствует, что устойчивые растения характеризуют более высокие содержание аскорбата и а-токоферола, активность антиоксидантных ферментов. В неблагоприятных условиях растения способны к индукции активности АОС. В результате они приобретают устойчивость не только к действующему агенту (например, гербициду), но и к другим воздействиям, вызывающим образование АФК. Такие факты послужили стимулом для создания с помощью методов генной инженерии форм растений, обладающих повышенной устойчивостью к гербицидам, инфекционным заболеваниям, загрязнителям атмосферы и др. Появились сообщения о создании организмов, обладающих очень высокой активностью СОД, аскорбатпероксидазы, глутатионредуктазы. Трансгенные растения с экспрессированной СОД оказывались более толерантными к водному дефициту, низкой температуре. Инфильтрация в ткани СОД подавляла как генерацию АФК, так и гибель пораженных патогеном клеток. Вместе с тем у растений, акклиматизированных к пониженной температуре, повышалась устойчивость к гербицидам, образующим АФК. Эти данные подтверждают рассмотренные выше представления о сопряженной устойчивости растений. Они свидетельствуют также в пользу гипотезы о том, что устойчивость растений к окислительному стрессу выступает важным фактором в приспособлении организмов к различным экстремальным воздействиям.
Уровень ферментативных АО находится под генетическим контролем. Повышение внутриклеточной концентрации O2 и H2O2 сопровождается активацией транскриптов генов антиоксидантов. Подобные эффекты усиления синтеза антиоксидантных ферментов выявлены при окислительном стрессе, индуцированном гербицидами. При ранении листьев зафиксирована активация экспрессии генов каталазы, и предполагается индуцирующее влияние H2O2. Трансгенные растения с повышенной экспрессией генов антиоксидантных ферментов, в частности СОД, характеризуются более высокой устойчивостью к абиотическим стрессам.
С окислительным стрессом связывают повреждения растений под действием ксенобиотиков, тяжелых металлов и алюминия, засоления, высоких интенсивностей света и ультрафиолетового облучения, засухи, дефицита элементов минерального питания, гипоксии и аноксии, загрязнителей атмосферы (SO2, H2S, CO, озона и др.), гипо- и гипертермии.
Защита растений от окислительного стресса. В последние годы активно обсуждаются следующие возможности направленного воздействия на устойчивость посредством усиления антиоксидантной защиты:
• прямая обработка природными и синтетическими антиоксидантами различных классов, например маннитом, защищающим хлоропластные ферменты от гидроксильного радикала, или селеном, защищающим растения от индуцированного УФ-В окислительного стресса;
• обработка аналогами фитогормонов и салициловой кислоты, обладающими антиоксидантными свойствами;
• действие оксидантов (H2O2, параквата) и индуцирующих экспрессию генов антиоксидантных ферментов повышает устойчивость к последующему окислительному стрессу;
• создание трансгенных растений, сверхэкспрессирующих чужеродные ферменты, повышающие устойчивость к окислительному стрессу (Mn-СОД в митохондриях и хлоропластах, глутатион-S-трансфераза, глутатионредуктаза и др.).
Все эти подходы, направленные на повышение уровня низкомолекулярных АО и активности антиоксидантных ферментов, в конечном счете приводят к усилению защиты от окислительного стресса и на этой основе — к более высокой резистентности растений к абиотическим и биотическим стрессорам.